
3 febrero de 2020
EL METABOLISMO
El metabolismo es un conjunto de reacciones que tiene lugar en el interior de la célula para satisfacer las necesidades de materia y energía de la misma.
La célula intercambia materia y energía con su entorno:
-
Sintetizando las sustancias necesarias para mantener las estructuras celulares en buen estado.
-
Realizando las funciones vitales.
Las reacciones metabólicas están relacionadas en las rutas o VÍAS METABÓLICAS. En estas intervienen las enzimas como mínimo una por reacción, de manera que el producto de una reacción es el sustrato de la siguiente.
En el metabolismo podemos diferenciar 2 tipos de rutas: catabolismo y anabolismo.
El CATABOLISMO es un conjunto de reacciones cuyo objetivo es la obtención de energía y moléculas precursoras sencillas mediante la degradación de moléculas complejas. Por tanto las características del catabolismo son:
-
Reacciones de oxidación y degradación.
-
Reacciones exergónicas, ya que se libera energía.
-
A partir de muchos sustratos se sintetizan casi siempre los mismos productos (CO2, ácido pirúvico, etanol).
-
Son vías metabólicas de convergencia.
El ANABOLISMO es un conjunto de reacciones químicas cuyo objetivo es la síntesis de moléculas complejas a partir de moléculas precursores sencillas. Por tanto la características del anabolismo son las siguientes:
-
Reacciones de síntesis y reducción.
-
Reacciones endergónicas, ya que se consume energía.
-
A partir de pocos sustratos se pueden obtener muchos productos.
-
Son vías metabólicas de divergencia.
Sus diferencias las encontramos reflejadas en este cuadro comparativo.
Ambos procesos están relacionados ya que la energía y las moléculas precursoras sencillas obtenidas en el catabolismo son necesarias para el desarrollo de las reacciones anabólicas.
Por tanto el OBJETIVO del metabolismo es: la obtención de energía, a través de la degradación de materia orgánica, la obtención de materia precursora sencilla necesaria para la síntesis de materia orgánica y por último la biosíntesis de moléculas orgánicas.
Antes de comenzar con el anabolismo es necesario comentar una serie de ideas, que explicaré a continuación.
Existen 2 tipos de organismos dependiendo de donde sacan la energía sus funciones: por un lado están los HETERÓTROFOS, su energía procede de sustancias elaboradas por otros seres vivos. Y por otro lado están los AUTÓTROFOS, que obtienen la energía de sustancias inorgánicas.
Partiendo de esa idea, el siguiente cuadro nos muestra los diferentes tipos de organismos:

Fuente: Creación propia

Fuente: Tema
Por otro lado, las reacciones químicas son transformaciones de energía, en las que la energía de unos enlaces se transfiere a otros nuevos. Cuando en estas reacciones se produce transferencia de electrones se llaman REACCIONES DE OXIDO-REDUCCIÓN, en las que cuando un compuesto se oxida (rompiendo enlaces con el hidrógeno y formando enlaces con el oxígeno), el otro se reduce (formando enlaces nuevos con el hidrógeno y rompiendo enlaces con el oxígeno).
Por otro lado, también es importante conocer que la energía se puede transportar de 2 maneras, por ATP o por coenzimas transportadoras de electrones, ambas ricoenergéticas. En el caso del ATP, podemos destacar los siguientes ideas:
es una COENZIMA DE TRANSFERENCIA, la cual se encarga de transportar grupos funcionales, actúa como MONEDA ENERGÉTICA y lo hace en todas las reacciones de intercambio de energía. Además está compuesto por adenina, tres fosfatos y ribosa (en esto es en lo que se parecen a los ácidos nucleicos, en la composición). Por lo que respecta a los enlaces de esos grupos fosfatos estos requieren de mucha energía, por lo que al sintetizar ATP a partir de ADP y Pi serán capaces de almacenar la energía, mientras que al descomponer el ATP en ADP y Pi esta energía podrá ser liberada.
De esta manera en las reacciones ENDERGÓNICAS (que son en las que se consume energía), lo que va a producirse es la hidrólisis del ATP dando lugar a ADP y Pi, de manera que debido a la ruptura de ese enlace se produce la obtención de 7 Kcal/mol. Sin embargo hay una ruta alternativa por la cual se puede obtener el doble de energía que consiste en la ruptura de los dos enlaces de alta energía del ATP, de manera que este se escinde en AMP y Pirofosfato, y posteriormente ese pirofosfato que son los dos fosfatos inorgánicos juntos se hidroliza. De esta forma podemos obtener el doble de energía es decir 14 Kcal/mol.
Mientras que en las reacciones EXERGÓNICAS (en las que se genera la energía), se sintetiza ATP a partir de ADP y Pi.
Se puede sintetizar de 3 formas que son:
FOTOFOSFORILACIÓN: es la síntesis de ATP que se produce en la membrana de los tilacoides. Lo que ocurre es que al incidir 2 fotones sobre el fotosistema ya sea I o II se liberan 2 electrones, estos pasarán por diferentes aceptores, dando lugar a la cadena transportadora de electrones. Por otro lado los electrones en el caso del fotosistema II perdidos se recuperan por la fotólisis de una molécula de agua. De esa manera lo que ocurre es que el transporte de electrones provoca el bombeo de protones por parte del citocromo B6-F del estroma al interior de los tilacoides, estos se juntan con los de la fotólisis de agua en caso de que haya fotosistemas II y generan una diferencia de potencial electroquímico entre las membranas, que hace que los protones salgan a través de la ATP-SINTETASA, produciéndose la síntesis de ATP.
FOSFORILACIÓN A NIVEL DE SUSTRATO: es la síntesis de ATP gracias a la energía que se libera en la degradación de biomoléculas, mediante la hidrólisis de algunos de sus enlaces ricos en energía. Este proceso es regulado por las QUINASAS.
FOSFORILACIÓN OXIDATIVA: es un proceso similar al anterior en este caso es la síntesis de ATP gracias a la energía liberada en el transporte de electrones en la cadena respiratoria de las mitocondrias. Este transporte provoca un bombeo de protones al espacio intermembrana, generándose una diferencia de gradiente electroquímico que impulsa los electrones de regreso a la matriz mitocondrial a través de la ATP-SINTETASA. De manera que la energía liberada por el flujo de H+ a favor de gradiente posibilita la unión de ADP y Pi.
Por lo que respecta al transporte de electrones (que en la mayoría de las reacciones transportan energía) lo llevan a cabo NADH, FADH2 Y NADPH, si se trata de electrones que forman hidrógenos que son desprendidos en las reacciones de oxidación. Mientras que si se trata de electrones libres desprendidos en las reacciones de oxidación, las moléculas que los captan son el citocromo y la coenzima A.
Por otro lado, cabe destacar la gran importancia que tienen las enzimas en el metabolismo celular.
Para que una reacción química se lleve a cabo es necesario que al sustrato se le suministre una cierta cantidad de energía, que se llama ENERGÍA DE ACTIVACIÓN, para llevarlo a un nivel de energía alto conocido como ESTADO DE TRANSICIÓN en el que los grupos funcionales se aproximan, se debilitan los enlaces y se puede producir la reacción. Las sustancias que se encargan de reducir la cantidad de energía de activación necesaria se llaman catalizadores. Las ENZIMAS son CATALIZADORES, más concretamente BIOCATALIZADORES ya que se encuentran en los seres vivos. Estos son proteínas o asociaciones de proteínas y otras moléculas orgánicas o inorgánicas, que actúan catalizando las reacciones químicas del metabolismo. Es decir, lo que hacen es acelerar y facilitar las reacciones al disminuir la cantidad de energía de activación necesaria para que estas se lleven a cabo.
Entre las características de las enzimas encontramos:
-
Aceleran y facilitan las reacciones.
-
No se consumen durante las reacciones.
-
Son eficaces en cantidades pequeñas.
-
Tienen una elevada especificidad.
-
Actúan siempre a la temperatura del organismo.
-
Tienen un elevado peso molecular.
-
Tienen en enorme poder catalítico.
En cuanto a su naturaleza se trata de proteínas globulares a excepción de unas moléculas de ARN que se llaman RIBOZIMAS que tienen función catalítica. Muchas enzimas son proteínas en el sentido estricto. Sin embargo hay enzimas que presentan una parte proteica APOENZIMA y una parte no proteica COFACTOR. Dentro de los cofactores pueden ser INORGÁNICOS, grupos prostéticos (si están unidos fuertemente a la apoenzima) u ORGÁNICOS, coenzimas (si están unidas débilmente a la apoenzima). La unión coenzima-apoenzima es temporal y similar a la enzima-sustrato, por ello a las enzimas se pueden considerar como un segundo sustrato. Estas coenzimas pueden ser COENZIMAS DE OXIDACIÓN Y REDUCCIÓN (NAD, NADP y FADH) que transportan electrones y protones, o COENZIMAS DE TRANSFERENCIA (ATP y Co-A) que transportan grupos funcionales.
Las VITAMINAS son precursores de coenzimas y de moléculas activas o son imprescindibles para su síntesis. Y hay 2 tipos: liposolubles (no son cofactores o precursores de coenzimas) e hidrosolubles (si que actúan como coenzimas o precursores).
En la parte PROTEICA enzimáticas podemos encontrar los siguientes aminoácidos:
-
ESTRUCTURALES: forman parte de la estructura pero no intervienen en el proceso catalítico.
-
DE FIJACIÓN: establecen uniones débiles con los sustratos. Constituyen el centro de fijación.
-
CATALIZADORES: se unen temporalmente al sustrato mediante enlaces covalentes fuertes, de manera que provocan cambios en los sustratos dando lugar a productos. Constituyen el centro catalítico.
El centro catalítico y el de fijación se encuentran próximos y constituyen lo que se conoce como CENTRO ACTIVO de la enzima que cuenta con las siguientes características:
-
Es una parte muy pequeña de la enzima.
-
Presenta una estructura tridimensional en forma de cavidad.
-
Presenta aminoácidos de fijación y catalizadores.
-
Algunos de sus aminoácidos presentan una gran afinidad por el sustrato.

Fuente: Tema
Para que las enzimas lleven a cabo su función primeramente debe actuar sobre ellas otra enzima o iones, que se llaman ZIMÓGENOS o PROENZIMAS.
En cuanto a su modo de actuación, la configuración de la parte proteica de la enzima es la que determinará la función de la misma. Para ello el sustrato se une temporalmente al centro activo de la enzima a través de los aminoácidos fijadores, formándose el complejo ENZIMA-SUSTRATO. Posteriormente los aminoácidos catalizadores del centro activo se unen débil o fuertemente al sustrato y provocan cambios en este, como el cambio en los grupos funcionales, dando lugar al complejo ENZIMA-PRODUCTO. A continuación el producto se separará de la enzima y obtendremos por un lado el producto y por otro lado la enzima intacta servible para otras reacciones. Esta actuación se lleva a cabo de formas distintos dependiendo de la cantidad de sustrato. Si se trata de un solo sustrato la enzima atrae al sustrato y lo une a su superficie produciéndose el proceso catalítico. Mientras que si son 2 sustratos pueden pasar 2 cosas, que ambos sean atraídos a la misma vez por la enzima, de esa forma el proceso catalítico se llevará a cabo a la mismo vez. Por otro lado, puede ocurrir que primero sea atraído uno de ellos, de manera que al desprenderse deja parte de el en el centro activo de la enzima y posteriormente el segundo se une a la parte fijada del primer sustrato.

Fuente: Tema
Por lo que respecta a la ESPECIFICIDAD de las enzimas, consiste en que solo si fijan a la enzima aquellas sustratos que puedan establecer enlaces con los aminoácidos de fijación del centro activo y de estas solo aquellas que presenten enlaces susceptibles de ruptura próximos a los aminoácidos catalizadores. De esa manera podemos encontrar varios modelos de especificidad: modelo de complementariedad, modelo de ajuste inducido y modelo de apretón de manos. Y por otro lado existen diferentes clases de especificidad: especificidad de grupo, de clase o absoluta.
Por último con respecto a las enzimas solo queda destacar la CINÉTICA ENZIMÁTICA que es el estudio de la velocidad a la que se producen las reacciones catalizadas por las enzimas. Esta velocidad a su vez depende de la concentración de sustrato, de enzima, la temperatura, el pH y la presencia o no de inhibidores. En esta se puede tratar de inhibición irreversible o reversible (dentro de la cual encontramos la competitiva, la acompetitiva y la no competitiva).
A continuación adjunto el esquema en el que aparece toda esta información:
El CATABOLISMO es un conjunto de reacciones del metabolismo que permite la degradación de moléculas complejas en moléculas más sencillas, liberándose energía. Existen varios tipos de rutas catabólicas:
-
GLUCÓLISIS: es el proceso en el que una molécula de glucosa (6C) se transforma en 2 moléculas de ácido pirúvico (3C).
-
B-OXIDACIÓN: es un conjunto de reacciones en las que se produce la oxidación de los ácidos grasos, dando lugar a un compuesto formado por 2 átomos de C que es el acetil-CoA.
-
TRANSAMINACIÓN: es un conjunto de reacciones que tiene lugar en el degradación de los aminoácidos, mediante la separación del grupo amino del esqueleto carbonado.
Las tres rutas convergen hacia la formación del acetil-CoA, que es el punto de partida del ciclo de krebs.
CATABOLISMO

Fuente: Creación propia

Fuente: https://slideplayer.es/
GLUCÓLISIS Y DESCARBOXILACIÓN OXIDATIVA
El CATABOLISMO DE LOS AZÚCARES se produce VÍA GLUCOSA, ya que es el más abundante de todo la naturaleza. Por tanto todas las reacciones convergen en una única ruta central de la degradación de la glucosa. De manera que los grandes polisacáridos de reserva mediante una reacción de fosforilación catalizada por la enzima glucógeno o almidón -fosforilasa se trnasforman en glucosa 1-fosfato, y posteriormente gracias a la enzima fosfomutasa se transforma a glucosa 6-fosfato que es primer intermediario de la glucólisis. Por otro lado los monosacáridos diferentes a la glucosa, se transformarán en esta o en alguno de los intermediarios. Así todos convergen hacia la misma ruta.
La primera etapa es la GLUCÓLISIS, en esta una molécula de glucosa (6C) se degrada para dar lugar a 2 moléculas de ácidos pirúvico (3C). En esta etapa se recupera parte de la energía contenida en la glucosa en forma de ATP y de NAH. Estas reacciones son anaerobias, ya que no se utiliza el oxígeno y se producen integramente en el citosol. Esta parece necesitar de una primer etapa de cebado para después poder liberar parte de la energía liberada en la glucosa en la segunda etapa, obteniendo así más energía. El proceso completo se encuentra en el esquema que adjuntaré posteriormente.
Algo a destacar en esta glucólisis es que tras haber obtenido la glucosa 6-fosfato podemos seguir otra RUTA que es la DE LAS PENTOSAS, cuyo objetivo no es el de obtener energía sino un azúcar de 5 átomos de carbono y NAPDH, necesarios para las reacciones del anabolismo. Lo que ocurre es que se producen 2 DESHIDROGENACIONES en las que la glucosa 6-fosfato pasa a 6-fosfoglucanato y posteriormente a ribulosa 5-fosfato. En ambas 1NADP+ se reduce a NADPH. Y por último se produce una DESCARBOXILACIÓN en la que la RIBULOSA 5-FOSFATO pasa a RIBOSA 5-FOSFATO.
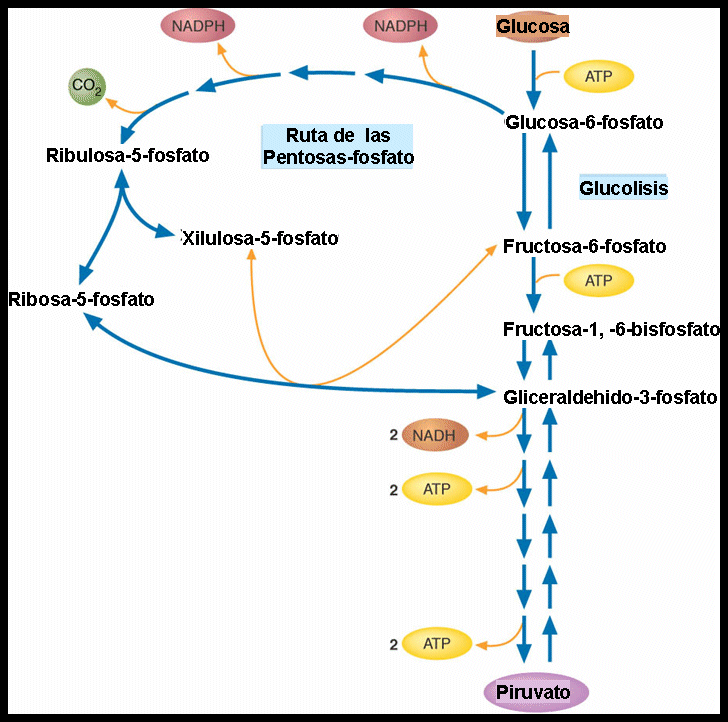
Fuente: https://slideplayer.es/
Una vez realizada la glucólisis el ácido pirúvico puede seguir 2 rutas, la de la respiración celular o la de las fermentaciones. La respiración celular es el proceso de degradación completa de la glucosa en presencia de oxígeno, para dar lugar a H2O y CO2, y a energía en forma de ATP. Si se sigue el camino de la respiración celular que se lleva a cabo en las mitocondrias, más concretamente en la matriz mitocondrial y en la membrana mitocondrial interna pasa lo siguiente. El ÁCIDO PIRÚVICO obtenido en la glucólisis penetra en la matriz mitocrondrial por las PERMEASAS de la membrana mitocondrial. Y una vez en el interior sufre una DESCARBOXILACIÓN OXIDATIVA, catalizada por el complejo de la PIRUVATO DESHIDROGENASA, dando lugar a Acetil-CoA y a CO2. Por cada molécula de ácido pirúvico que entra se consumen 2 NADH y un Coenzima-A. Así se obtienen 2 moléculas de acetil-CoA.

Fuente: https://slideplayer.es/
CICLO DE KREBS
El acetil-CoA obtenido puede oxidarse completamente en la misma mitocondria por el CICLO DE KREBS, CICLO DEL ÁCIDO CÍTRICO O CICLO DE LOS ÁCIDOS TRICARBOXÍLICOS. Este se inicia con la condensación del acetil-CoA (2C), con el ácido oxalacético (4C), dando lugar a un compuesto formado por 6C que es el ácido cítrico. En este proceso también se libera un coenzima A. A continuación se produce una secuencia de 7 reacciones consecutivas en las que se desprende carbono en forma de CO2 y se regenera el ácido oxalacético.
Primero el ácido cítrico pasa a su isómero isocítrico desprendiéndose una molécula de H2O. Posteriormente el ISOCÍTRICO se transforma en ÁCIDO ALFA-CETOGLUTÁRICO (aparece una cetona), reduciéndose el NAD+ a NADH, y desprendiéndose carbono en forma de CO2, es decir se produce una DESCARBOXILACIÓN OXIDATIVA. A continuación, este compuesto vuelve a sufrir una descarboxilación, reduciéndose el NAD+ a NADH y desprendiéndose CO2, así se obtiene el SUCCINIL-CoA. Este pasa en una fosforilación a nivel de sustrato a ÁCIDO SUCCINICO, formándose GTP, análogo al ATP. Posteriormente este compuesto pasa a ÁCIDO FUMÁRICO, mediante una OXIDACIÓN en la que se reduce el FADH a FADH2. El ácido fumárico pasa a ÁCIDO MÁLICO mediante la incorporación de una molécula de H2O. Y finalmente este pasa mediante una OXIDACIÓN a ácido oxalacético otra vez, reduciéndose en este proceso el NAD+ a NADH. De manera que por cada vuelta se obtienen 3NADH, 2FADH2 y 2CO2. De esta manera se completa la degradación del esqueleto carbonado de la glucosa que poseía 6 carbonos, pues 2 de ellos han sido perdidos en la descarboxilación oxidativa, y como son 2 acetil-CoA los que tenemos se dan 2 vueltas al ciclo de krebs por lo que se desprenden 4 carbonos que sumados a los 2 de la fase anterior conforman los 6 C de la glucosa.

Fuente: Tema
CADENA RESPIRATORIA
Sin embargo, el rendimiento energético continúa siendo escaso, solo se obtienen 2 ATP en la glucólisis y 2 en el ciclo de krebs. Y es que parte de la energía de la glucosa se encuentra retenida en los electrones de alta energía de los coenzimas reducidos NADH y FADH2 obtenidos en las anteriores etapas. Esta energía será liberada en la cadena de transporte de electrones cuando el NADH y el FADH2 cedan sus electrones a una de las miles de cadenas de transporte electrónico que hay distribuidas por la membrana interna de las mitocondrias, los cuales van pasar a través de los diferentes complejos que componen a la cadena, que son 4 grandes complejos proteicos, una molécula lipídica pequeña (UBIQUINONA) y una molécula proteica pequeña (CITOCROMO C). Esta cadena va a transportar los electrones hasta el OXÍGENO que es el último aceptor ÚLTIMO ACEPTOR de electrones, el cual se reduce para dar lugar a H2O que es uno de los productos finales de la degradación de la glucosa. Ese transporte de electrones se produce mediante una secuencia de reacciones redox, en las que intervienen 2 transportadores uno cede un par de electrones reduciéndose y el otro los capta oxidándose. Esos transportadores a su vez no están colocados de manera irregular sino que se colocan de menor a mayor potencial redox. Por tanto el transporte de electrones se produce a favor de diferencia de POTENCIAL REDOX (proceso fuertemente EXERGÓNICO, en el que se libera una gran cantidad de energía).
El NADH cede sus electrones al complejo I, bombeando 10 H+ al espacio intercelular. Sin embargo el FADH2 los cede a la ubiquinona por lo que eluden el complejo I, y por tanto solo bombean 6 H+. Hay tener en cuenta que el NADH obtenido en el citosol tendrá que pasar al interior de la mitocondria a través de un SISTEMA DE LANZADERA.
La energía que se libera en este transporte es aprovechada por la célula mediante el acoplamiento de una fosforilación oxidativa al transporte electrónico.
El proceso por tanto consiste en que primeramente el transporte de electrones provoca el bombeo de protones de la matriz mitocondrial al espacio intermembrana (QUIMIÓSMOSIS) y este bombeo genera un gradiente electroquímico (en el que se almacena la energía liberada en el transporte de electrones). De esa forma los protones van a tender a entrar otra vez en la matriz mitocondrial a tráves de las ATP-SINTETASA que se encuentran en la cadena respiratoria. Esta entrada se va a producir a favor de gradiente electroquímico. Por lo que gracias al flujo de protones a través de la ATP-SINTETASA a favor de gradiente electroquímico se va a liberar la energía que va a fosforilar el ADP y Pi a ATP. (FOSFORILACIÓN OXIDATIVA)

Fuente: Tema
FERMENTACIONES
Sin embargo, también podemos seguir el camino de las fermentaciones. La fermentación es un proceso catabólico en el que no hay cadena de transporte electrónico como hay en la respiración celular. De esa manera no hay tampoco hay ATP-SINTETASA, lo que explica porque siempre se produce una FOSFORILACIÓN A NIVEL DE SUSTRATO, lo que a su vez explica el escaso rendimiento energético. Además el último aceptor no es el oxígeno ya que la fermentación es un proceso ANAEROBIO, sino que es una molécula orgánica.
Esta como ha sido mencionado anteriormente, se produce en microorganismos como las levaduras o las bacterias, pero también en las células musculares, cuando tras haberse realizado un ejercicio intenso no se ha producido la llegada de suficiente oxígeno por la sangre.
El verdadero significado de la fermentación está en la siguiente idea. Cuando una molécula de glucosa se degrada en la glucólisis da lugar a dos moléculas de NADH, pero ese NADH en la célula es limitado por lo que deberá reciclarse para que el proceso no sea parado. En su caso las células aerobias lo que hacen es reducir el NADH cediendo este sus electrones al oxígeno, que es el último aceptor de electrones. Mientras que en las células anaerobias, el NADH cede sus electrones a una molécula orgánica que puede ser el ácido pirúvico o algún producto de la degradación de este, mediante un o varias reacciones adicionales a la glucólisis. A esas reacciones adicionales se les llama fermentación y por tanto el principal objetivo de esta es el reciclaje del NADH.
La FERMENTACIÓN consiste en la transformación del ácido pirúvico en productos más sencillos que son diferentes en función del tipo de fermentación que se de, y es que existen varios tipos de fermentaciones que proporcionan una amplia gama de productos con intereses industriales y alimenticios.
Un tipo de fermentación es la HOMOLÁCTICA, que solo tiene una etapa en la que el ácido pirúvico acepta un par de electrones del NADH y se reduce a ácido láctico que es el producto final. Este tipo de fermentaciones se da en bacterias como el Lactobacillus, y se obtienen productos derivados de la leche, como queso, yogurt y requesón, de ahí el interés de esta fermentación en la industria alimenticia. Por otro lado encontramos la fermentación heteroláctico cuyo producto es mixto (ácido láctico, alcohol etílico y CO2).
Otro tipo de fermentación es la ALCOHÓLICA, esta consta de 2 etapa, en la primera el ácido pirúvico pierde el grupo carboxilo en forma de CO2, convirtiéndose en ACETALDEHIDO y en la segunda el acetaldehido acepta un par de electrones del NADH transformándose en ALCOHOL ETÍLICO que es el producto final. Pero existen aún más fermentaciones como la BUTÍRICA y la PUTRIDA, en el caso de la butírica, se produce la descomposición de sustancias glucídicas de origen vegetal, almidón, celulosa,... en ácido butírico, CO2, hidrógeno y sustancias malolientes. Y en el caso de la putrida es la descomposición pero de sustancias de naturaleza proteica o aminoacídica, dando lugar a sustancias orgánicas y malolientes, algunos de los productos que son poco desagradables de esta fermentación se utilizan para producir los sabores típicos del queso, del vino,...
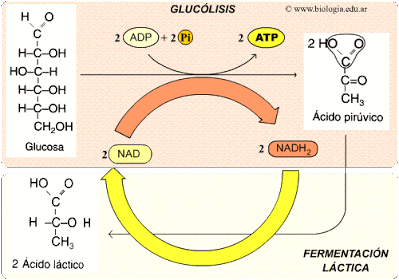

Fuente: Tema
CATABOLISMO DE GLÚCIDOS, LÍPIDOS Y PROTEÍNAS
Además de los azúcares, también hemos estudiado el catabolismo de lípidos, proteínas y ácidos nucleicos.
En el caso de los lípidos, generalmente se trata de TRIGLICÉRIDOS ya que son los lípidos que más energía aportan. Estos comienzan su degradación con la separación de la GLICERINA y de los 3 ÁCIDOS GRASOS gracias a la enzima LIPASA. Y una vez obtenidos la glicerina y los ácidos grasos, por un lado se producirá la degradación de la glicerina y por otro la de los ácidos grasos.
Por lo que respecta a la oxidación de la GLICERINA esta se fosforila pasando a glicerol 3-fosfato y consumiéndose ATP. Y a continuación el glicerol 3-fosfato se oxida dando lugar a dihidroxiacetona 3-fosfato y a NADH.
Por otra lado, los ÁCIDOS GRASOS, estos van a penetrar en la matriz mitoncondrial donde se va a llevar a cabo la B-oxidación de estos. Para ello primeramente deben ser activados por el coenzima-A transformándose en Acil(graso)-CoA. Esta activación tiene lugar cuando el ácido graso atraviesa la membrana mitocondrial externa y requiere de ATP, de manera que este se hidroliza para dar AMP y pirofosfato, que posteriormente se dividirá en dos Pi, así se han consumido dos enlaces fosfato alto energéticos. Por lo que aquí ya tenemos el consumo de una molécula de ATP y un Coenzima-A para activar al ácido graso. (solo se activa una vez, es decir que cuando se produce la héliza de Lynen no tiene que volver a activarse conforme vaya completando la vuelta porque ya lo está)
Una vez activado ya se produce la vuelta a la HÉLICE DE LYNEN, en esta primeramente aparece un doble enlace entre el carbono alfa y beta, en este proceso el FADH se reduce a FADH2 (una de las moléculas que se libera). Posteriormente y mediante la incorporación de una molécula de H2O (otra de las moléculas que se consume), el OH de esta se une al carbono beta. Y a continuación ese OH se transforma en un grupo cetona, en este proceso se produce la reducción del NAD a NADH (otra de las moléculas que se libera). Y por último el acil-CoA sufre la pérdida del Acetil-CoA (otra de las moléculas que se libera). En ese momento obtenemos un ácido graso con 2 átomos de carbono menos y este continúa realizando tantas vueltas como sea necesario hasta que la oxidación del ácido graso sea completa. De manera que si se trata de un ácido graso de 14 átomos de carbono se obtendrán 14 acetil- CoA y se darán 13 vueltas, por lo que se obtendrán 13 FADH2 y 13 NADH.

Fuente: Tema
Por otro lado las PROTEÍNAS no se suelen utilizar como combustibles metabólicos, solo en caso de extrema necesidad. En ese caso se separarán en sus AMINOÁCIDOS, gracias a las proteasas. Y ya obtenidos los aminácidos (si son sobrantes) se llevará a cabo la separación del grupo amino (TRANSAMINACIÓN y DESAMINACIÓN OXIDATIVA) y por otro lado la DEGRADACIÓN DEL ESQUELETO CARBONADO.
Por lo que respecta a los ÁCIDOS NUCLEICOS tampoco suelen utilizarse como combustible metabólico, pero si eso ocurre se separan en sus NUCLEÓTIDOS (gracias a las nucleasas), los cuales si son sobrantes se procederá a degradar en sus componentes (bases nitrogenadas, pentosas y ácido fosfórico).
A continuación os adjunto el esquema general del catabolismo:
ANABOLISMO
El ANABOLISMO es la vía constructiva del metabolismo, en la que a partir de moléculas sencillas y energía se da lugar a moléculas más complejas. Si la molécula inicial es orgánica se trata de anabolismo heterótrofo, este se da en todos los organismos. Y si la molécula inicial es inorgánica se trata de anabolismo autótrofo. Este a su vez se divide en:
-
FOTOSÍNTESIS: es el anabolismo que se produce gracias a la captación de energía luminosa.
-
QUIMIOSÍNTESIS: es el anabolismo que se produce gracias a la energía desprendida en reacciones de oxidación de moléculas.
En primer lugar la FOTOSÍNTESIS consiste en la transformación de energía luminosa en energía química que primeramente es almacenada por el ATP pero luego es utilizada por la fase oscura de la fotosíntesis es decir para la obtención de materia orgánica. Hay dos tipos de fotosíntesis:
-
OXIGÉNICA: en la que el dador de electrones es el H2O, por tanto se desprende oxígeno.
-
ANOXIGÉNICA: el dador de electrones no es el H2O, sino otra sustancia como el sulfuro de hidrógeno, por ello no se desprende energía.
La fotosíntesis consta de 2 etapas:
-
FASE LUMINOSA: es la etapa en la que se produce la captación de la energía luminosa por parte de los pigmentos fotosintéticos de la membrana de los tilacoides de los cloroplastos. En esta se genera ATP y NADPH.
-
FASE OSCURA O CICLO DE CALVIN: no requiere de luz y es el proceso en el que se produce la fijación del CO2 atmosférico. En esta etapa se utilizan el NADPH y el ATP obtenidos en la fase anterior.
En primer lugar encontramos la fase luminosa, en la cual se capta la energía luminosa gracias a los pigmentos fotosintéticos de la membrana de los tilacoides de los cloroplastos. En esta fase se obtienen el ATP y el NADPH que serán utilizados en la siguiente fase, que es la fase oscura, para la síntesis de materia orgánica.
En la fase luminosa cíclica lo que va a ocurrir es que sobre el fotosistema II (que es el primero que interviene) van a incidir 2 fotones que van a excitar su pigmento diana que es la clorofila P680 del centro de reacción. Esta va a perder 2 electrones que recuperará gracias a la hidrólisis de la molécula de agua, que se llama fotólisis del agua, es decir la ruptura de los enlaces del agua, en la que se obtendrá 2H+, 2 electrones y 1/2 de O2. Esos 2 electrones perdidos son captados por el aceptor primario que es la feofitina, luego pasan a la PLASTOQUINONA, esta los cede al citocromo b6-f (que bombea protones del estroma al espacio tilacoidal) y de este pasan a la PLASTOCIANINA. Finalmente son captados por la clorofila P700 del fotosistema I. De esta manera los 2 electrones que la clorofila P700 del fotosistema I ha perdido al incidir sobre el 2 fotones son recuperados. Los 2 electrones perdidos por el fotosistema I pasan a la FERREDOXINA, que los transporta hasta la NADP+ REDUCTASA, donde van a llegar 2H+ del estroma para formar hidrógeno que será captado por el NADP+ para reducirse a NADPH+ (fotorreducción del NADP+).
Mientras que con los H+ lo que va a ocurrir es lo siguiente. La energía desprendida en el transporte de electrones va bombear H+ del estroma al interior del tilacoide (por el citocromo b6-f) y estos junto con los 2 de la fotólisis del agua van a generar una diferencia de potencial electroquímico a ambos lados de la membrana que va a hacer que los protones salgan al estroma a través de la ATP-SINTETASA, produciéndose la fotofosforilación, es decir la formación de ATP.
De esta manera hemos obtenido a partir de una molécula de agua y 4 fotones, 1 NADPH, 1/2 O2 y un ATP. (Por cada 2 electrones entran 4 H+, 2 de la hidrólisis del agua y 2 impulsados por la cadena de transporte,
y tres protones dan lugar a 1 ATP). Sin embargo se requiere de 3 ATP para que se produzca la fase oscura por lo que se deberá llevar a cabo una fase luminosa cíclica.
En la fase luminosa cíclica solo actúa el fotosistema I, cuya clorofila P700 pierde 2 electrones por la incidencia de 2 fotones que volverán otra vez a el, por lo que no hay ni fotólisis del agua, reducción del NADP+, ni se desprende O2. En este caso inciden 2 fotones sobre el fotosistema I que excitan a la clorofila P700, de manera que pierde 2 electrones, estos pasan a la FERREDOXINA, después al CITOCROMO B6-F, (que bombea los protones desde el estroma al interior de los tilacoides, y cuando estos salen por la ATP-SINTETASA, provocan la síntesis de ATP), posteriormente pasan a la PLASTOCIANINA y luego al PSI otra vez. De manera que así hemos obtenido la cantidad de energía que necesitábamos para realizar la fase oscura.
Por tanto el balance de esta fase luminosa es el siguiente: En primer lugar, en la fase acíclica
a partir de una molécula de agua y 4 fotones, obtenemos 1 NADPH, 1/2 O2 y un ATP. (Por cada 2 electrones entran 4 H+, 2 de la hidrólisis del agua y 2 impulsados por la cadena de transporte,
y tres protones dan lugar a 1 ATP). Mientras que en la fase luminosa cíclica solo se produce ATP, pero es muy importante ya que en la fase oscura se requiere de más ATP del que se puede producir en la fase luminosa acíclica. Ese ATP y NADPH obtenidos se utilizan en la fase oscura.
FOTOSÍNTESIS

Fuente: Creación propia


Fuente: Tema
En la fase oscura de la fotosíntesis en la que no se requiere de luz, lo que se produce es la síntesis de materia orgánica a partir de CO2, utilizándose el ATP y el NADPH obtenido en la fase luminosa. Esta fase se da lugar en el estroma de los cloroplastos. A grandes rasgos se podría decir que es la fijación del carbono a partir de CO2, para dar lugar a una molécula orgánica más estable. Este se divide en tres fases que son la fijación del CO2, la reducción del CO2 fijado y la regeneración de la ribulosa 1,5-difosfato, las cuales se llevan a cabo de la siguiente manera:
Primeramente la pentosa ribulosa 1,5-difosfato se une en el estroma al CO2 atmosférico gracias a la enzima ribulosa 1,5-difosfato carboxilasa oxidasa, conocida con el nombre de ribusco, que es la que posibilita esa fijación del CO2 en la pentosa. De esa manera se da lugar a un compuesto inestable de 6 átomos de carbono el cual se divide en 2 moléculas de ácido 3-fosfoglicérico, que son moléculas de 3 átomos de carbono.
Posteriormente, se produce la reducción de ese CO2 fijado. En este caso, primero el ácido 3-fosfoglicérico es fosforilado, consumiéndose de esa manera ATP (obtenido en la fase luminosa, que se reduce a ADP y Pi) y así obtenemos 1,3-difosfoglicérico. Y a continuación este se reduce consumiéndose el NADPH (obtenido en la fase luminosa que se reduce a NADP+) y así obtenemos gliceraldehido 3-fosfato. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (puede quedarse en el estroma para la síntesis de almidón, ácidos grasos, aminoácidos,.. o puede ir al citosol para dar lugar a la glucosa o fructosa).
En el caso de la regeneración de la ribulosa 1,5-difosfato, lo que ocurre es que el gliceraldehido 3-fosfato se transforma en ribulosa-5-fosfato mediante un proceso complejo en el que se suceden una serie de compuestos de 3,4,5 y 7 carbonos. Al final se regenera la ribulosa 1,5-difosfato mediante la fosforilación directa con ATP. (este proceso es conocido como ciclo de las pentosas fosfato)
Por tanto en el ciclo de calvin por cada CO2 que se incorpora se requiere de 2 NADPH y 3 ATP. Este es un proceso que no requiere de energía luminosa ni de clorofila, pero se suele llevar a cabo durante el día porque es cuando se sintetizan el ATP y el NADPH.

Fuente: Tema
QUIMIÓSMOSIS
Por lo que respecta a la QUIMIÓSMOSIS, consiste en la síntesis de ATP a partir de la energía que se desprende en las reacciones de oxidación de ciertas sustancias inorgánicas. Y posteriormente este ATP se utiliza para convertir la materia inorgánica en materia orgánica. En esta también diferenciamos dos fases, una en la que se produce la oxidación de la sustancia inorgánica con la que se obtiene la energía y la otra en la que se produce la incorporación del CO2 en el ciclo de calvin a la materia inorgánica utilizándose la energía obtenida en la anterior fase, como pasaba en la fotosíntesis.
ANABOLISMO HETERÓTROFO
Por otro lado encontramos el anabolismo heterótrofo, este se da en todos los organismos y la molécula inicial que se toma es una molécula orgánica.
Este es el caso del anabolismo de glúcidos, el cual se divide en 2 fases: obtención de la glucosa (por GLUCONEOGÉNESIS o por el ciclo de calvin) y obtención de polímeros de glucosa (AMILOGÉNESIS o GLUCOGENOGÉNESIS).
También encontramos el caso del anabolismo de lípidos en el que primeramente debemos obtener la glicerina y los ácidos grasos, dando el MALONIL-COA desempeña un papel muy importante, para luego poder unirlos por esterificación.
Además de estos dos casos, también encontramos el caso del anabolismo de las proteínas en el que primero deberemos obtener aminoácidos. Y por último encontramos el anabolismo de ácidos nucleicos, en el que primero deberemos obtener los nucleótidos.
A continuación adjunto el esquema general del anabolismo por un lado el la fotosíntesis y quimiosíntesis y por otro lado el de anabolismo heterótrofo.


Fuente: Creación propia
Fuente: Creación propia
ACTIVIDADES
1.- ¿Cómo y cuándo tiene lugar la descomposición del agua en el proceso de fotosíntesis? ¿Cuáles son sus consecuencias?
Lo que ocurre es que en la primera fase de la fotosíntesis que es la fase luminosa, se produce la captación de energía luminosa por parte de los pigmentos del complejo captador de luz o centro antena de los fotosistemas. De esa manera esos pigmentos lo que van a hacer es transportar esa energía hasta el centro de reacción donde está el pigmento diana (clorofila a especial), y posteriormente esa energía va a ser captada por un electrón de la clorofila a y este salta y se dirige a un aceptor de electrones y posteriormente a otros aceptores dando lugar a la cadena de transporte de electrones. Sin embargo lo que le ha ocurrido a la clorofila a es que ha perdido un electrón que necesitará recuperar de algún modo, y la manera de recuperarlo es mediante la hidrólisis de la molécula de agua, que se llama fotólisis.
Este proceso se produce en la fase luminosa acíclica en la que intervienen los dos tipos de fotosistemas, el I y el II, mientras que en la fase luminosa cíclica (en la que solo participa el fotosistema I) cuando inciden 2 fotones sobre el fotosistema, la clorofila de este que es la P700, va a perder 2 electrones pero esos 2 electrones van a volver posteriormente ya que se trata de una fase cíclica.
Así en la fase luminosa acíclica lo que va a ocurrir es que sobre el fotosistema II (que es el primero que interviene) van a incidir 2 fotones de manera que la clorofila P680 del centro de reacción va a perder 2 electrones que recuperará gracias a la hidrólisis de la molécula de agua, que se llama fotólisis del agua, es decir la ruptura de los enlaces del agua. Esta fotólisis da lugar a 2 protones, 2 electrones y 1/2 de O2. Esos electrones se transportarán por la cadena transportadora de electrones pasando por el fotosistema I (por lo que cuando sobre este inciden 2 fotones, no necesita la ruptura de más agua para recuperar los 2 electrones sino que los recibe de la cadena de transporte). Posteriormente los electrones perdidos en el fotosistema I continúan su transporte hasta la NADP+ REDUCTASA, donde se incorporan dos H+ del estroma para formar hidrógeno y este es recogido por el NADP+ para reducirse a NADPH (fotorreducción del NADP+), que posteriormente será utilizado en el ciclo de calvin. Mientras que con los H+ lo que va a ocurrir es lo siguiente. La energía desprendida en el transporte de electrones va bombear H+ del estroma al interior del tilacoide (por el citocromo b6-f) y estos junto con los 2 de la fotólisis del agua van a generar una diferencia de potencial electroquímico a ambos lados de la membrana que va a hacer que los protones salgan al estroma a través de la ATP-SINTETASA, produciéndose la fotofosforilación, es decir la formación de ATP.

2.- Cloroplastos y fotosíntesis.
A) Durante el proceso fotosintético, coexisten un flujo cíclico y un flujo no cíclico de electrones. Exponga brevemente el sentido fisiológico de cada uno de ellos y cuáles son sus componentes principales.
En la fase luminosa acíclica lo que va a ocurrir es que sobre el fotosistema II (que es el primero que interviene) van a incidir 2 fotones de manera que se va a excitar su pigmento diana que es la clorofila P680 del centro de reacción. Esta va a perder 2 electrones que recuperará gracias a la hidrólisis de la molécula de agua, que se llama fotólisis del agua, es decir la ruptura de los enlaces del agua, en la que se obtendrán 2H+, 2 electrones y 1/2 de O2. Esos 2 electrones perdidos son captados por el aceptor primario que es la feofitina, luego pasan a la PLASTOQUINONA, esta los cede al citocromo b6-f (que bombea protones del estroma al espacio tilacoidal) y de este pasan a la PLASTOCIANINA. Finalmente son captados por la clorofila P700 del fotosistema I. De esta manera los 2 electrones que la clorofila P700 del fotosistema I ha perdido al incidir sobre el 2 fotones son recuperados. Los 2 electrones perdidos por el fotosistema I pasan a la FERREDOXINA, que los transporta hasta la NADP+ REDUCTASA, donde van a llegar 2H+ del estroma para formar hidrógeno que será captado por el NADP+ para reducirse a NADPH+ (fotorreducción del NADP+).
Mientras que con los H+ lo que va a ocurrir es lo siguiente. La energía desprendida en el transporte de electrones va bombear H+ del estroma al interior del tilacoide (por el citocromo b6-f) y estos junto con los 2 de la fotólisis del agua van a generar una diferencia de potencial electroquímico a ambos lados de la membrana que va a hacer que los protones salgan al estroma a través de la ATP-SINTETASA, produciéndose la fotofosforilación, es decir la formación de ATP.
De esta manera hemos obtenido a partir de una molécula de agua y 4 fotones, 1 NADPH, 1/2 O2 y un ATP. (Por cada 2 electrones entran 4 H+ y tres protones dan lugar a 1 ATP). Sin embargo se requiere de 3 ATP para que se produzca la fase oscura por lo que se deberá llevar a cabo una fase luminosa cíclica. En esta solo actúa el fotosistema I, cuya clorofila P700 pierde 2 electrones por la incidencia de 2 fotones que volverán otra vez a el, por lo que no hay ni fotólisis del agua, ni reducción del NADP+, ni se deprende O2. En este caso inciden 2 fotones sobre el fotosistema I que excitan a la clorofila P700, de manera que pierde 2 electrones, estos pasan a la FERREDOXINA, después al CITOCROMO B6-F, (que bombea los protones desde el estroma al interior de los tilacoides, y cuando estos salen por la ATP-SINTETASA, provocan la síntesis de ATP), posteriormente pasan a la PLASTOCIANINA y luego al PSI otra vez. De manera que así hemos obtenido la cantidad de energía que necesitábamos para realizar la fase oscura.
B) Existen algas procarióticas (cianobacterias) que carecen de cloroplastos y sin embargo realizan el proceso fotosintético de forma similar a como lo realizan las plantas superiores. ¿Cómo es posible?
Esto se debe a que aunque carezcan de cloroplastos presentan tilacoides en el citoplasma con los pigmentos fotosintéticos. De manera que cuenta con todos los lugares necesarios para que se pueda producir la fotosíntesis. Membrana de los tilacoides (fase luminosa) y estroma (ciclo de calvin).
3.- Explique brevemente la finalidad que tienen los siguientes procesos: - metabolismo - Respiración celular - Anabolismo - Fotosíntesis - Catabolismo
METABOLISMO: es el conjunto de reacciones químicas que tienen lugar en el interior de las células para satisfacer las necesidades de materia y energía de la misma. Todas estas reacciones se relacionan en unas rutas o vías metabólicas, en las que en cada reacción interviene como mínimo una enzima, de manera que el producto de una reacción es el sustrato de la siguiente. Hay 2 tipos de rutas el anabolismo y el catabolismo. Y el objetivo del metabolismo es la obtención de energía, a través de la degradación de materia orgánica, la obtención de materia precursora sencilla necesaria para la síntesis de materia orgánica y por último la biosíntesis de moléculas orgánicas.
RESPIRACIÓN CELULAR: es el proceso de la degradación completa de la glucosa en presencia de oxígeno. (camino que se sigue tras haberse realizado la glucólisis). Por tanto la finalidad del mismo es la degradación de la molécula de glucosa, para dar lugar a H2O, CO2 y energía en forma de ATP.
ANABOLISMO: es una vía constructiva del metabolismo, es decir, es la ruta en la que se produce la síntesis moléculas complejas a partir de moléculas sencillas y energía. Por tanto son reacciones de síntesis, de reducción y endergónicas (se consume energía). Además en estas a partir de unos pocos sustratos se obtiene una gran variedad de productos. Y por otro lado son vías divergentes.
FOTOSÍNTESIS: es un proceso anabólico en el que la energía luminosa (captada gracias a los pigmentos fotosintéticos de la membrana de los tilacoides de los cloroplastos) se transforma en energía química, que se almacena primeramente en forma de ATP para luego ser utilizada para sintetizar moléculas orgánicas más estables. Por tanto lo que ocurre es que en el ciclo de calvin se fija el CO2 en sustancias orgánicas utilizando el ATP y el NADPH obtenidos en la fase luminosa, de manera que obtenemos una molécula orgánica más compleja.
CATABOLISMO: es un conjunto de reacciones del metabolismo cuyo objetivo es la obtención de moléculas precursoras sencillas y energía, a partir de la degradación de biomoléculas. Por tanto son reacciones de degradación, de oxidación y exergónicas (se libera energía). Además en estas a partir de muchos sustratos se obtienen casi siempre los mismos productos (etanol, ácido pirúvico, CO2). Y se trata de vías convergentes.
4.- Defina: Fotosíntesis, fotofosforilación, fosforilación oxidativa y quimiosíntesis.
FOTOSÍNTESIS: es un proceso anabólico, más concretamente de anabolismo autótrofo, en el que la energía luminosa se transforma en energía química, que se almacena primeramente en forma de ATP para luego ser utilizada para sintetizar moléculas orgánicas más estables. Por tanto lo que ocurre es que en el ciclo de calvin se fija el CO2 en sustancias orgánicas utilizando del ATP y el NADPH obtenidos en la fase luminosa, obteniéndose una molécula orgánica más compleja. De esa manera establecemos que la fotosíntesis tiene dos fases, una luminosa (cíclica o acíclica) y una oscura (ciclo de calvin). Y por otro lado que hay dos tipos de fotosíntesis, la oxigénica en la que el dador de electrones es el H2O y se desprende oxígeno, y la anoxigénica en la que el dador de electrones es el sulfuro de hidrógeno y no se desprende oxígeno. Esta se da en algas, cianobacterias, plantas y bacterias fotosintéticas.
FOTOFOSFORILACIÓN: es la síntesis de ATP que se produce en la membrana de los tilacoides. Lo que ocurre es que al incidir 2 fotones sobre el fotosistema ya sea I o II se liberan 2 electrones, estos pasarán por diferentes aceptores, dando lugar a la cadena transportadora de electrones. Por otro lado los electrones en el caso del fotosistema II perdidos se recuperan por la fotólisis de una molécula de agua. De esa manera lo que ocurre es que el transporte de electrones provoca el bombeo de protones por parte del citocromo B6-F del estroma al interior de los tilacoides, estos H+ se juntan con los de la fotólisis de agua en caso de que haya fotosistema II y generan una diferencia de potencial electroquímico entre las membranas, que hace que los protones salgan a través de la ATP-SINTETASA, produciéndose la síntesis de ATP, al aprovecharse la fuerza protón motriz de vuelta de los protones.
FOSFORILACIÓN OXIDATIVA: es un proceso similar al anterior en este caso es la síntesis de ATP gracias a la energía liberada en el transporte de electrones en la cadena respiratoria de las mitocondrias. Este transporte provoca un bombeo de protones al espacio intermembrana, generándose una diferencia de gradiente electroquímico que impulsa los electrones de regreso a la matriz mitocondrial a través de la ATP-SINTETASA. De manera que la energía liberada por el flujo de H+ a favor de gradiente posibilita la unión de ADP y Pi.
QUIMIOSÍNTESIS: es un proceso anabólico, más concetramente es un proceso de anabolismo autótrofo, que consiste en la síntesis de ATP a partir de la energía que se desprende en las reacciones de oxidación de ciertas sustancias inorgánicas. Y posteriormente este ATP se utiliza para convertir la materia inorgánica en materia orgánica. En esta también diferenciamos dos fases, una en la que se produce la oxidación de la sustancia inorgánica con la que se obtiene la energía y la otra en la que se produce la incorporación del CO2 en el ciclo de calvin a la materia inorgánica utilizándose la energía obtenida en la anterior fase, como ocurría en la fotosíntesis.
5.- Anabolismo y catabolismo. Citar dos ejemplos de cada uno de estos procesos y en qué orgánulos celulares se producen.
-
Catabolismo: respiración celular (en las mitocondrias) y fermentaciones (en el citosol)
-
Anabolismo: fotosíntesis (cloroplastos) y quimiosíntesis (en bacterias quimioautótrofas).
6.- Un proceso celular en eucariota genera ATP y NADPH (H) con producción de oxígeno por acción de la luz sobre los pigmentos. ¿De qué proceso se trata? ¿Para qué se utiliza el ATP y el NADPH formados? ¿Participan los cloroplastos (indicar brevemente cómo).
Se trata de la fase luminosa acíclica de la fotosíntesis que se produce en la membrana de los tilacoides de los cloroplastos (por tanto los cloroplastos si que participan, ya que la fase luminosa se da en la membrana de los tilacoides y la fase oscura en el estroma). En esta, cuando 2 fotones inciden sobre el fotosistema II excitan el pigmento diana de este, de manera que la clorofila P680 pierde 2 electrones que se irán transportando por aceptores hasta llegar al fotosistema I donde como también inciden 2 fotones la clorofila P700 pierde 2 electrones, que recupera de los 2 que recibe de la cadena. Estos 2 electrones perdidos se transportarán hasta llegar a la NADP+ REDUCTASA en la que se reducirá el NADP+ a NADPH. Mientras que por otro lado en el fotosistema II la pérdida de esos 2 electrones se ha compensado con la llegada de otros 2 procedentes de la fotólisis del H2O. Además esta fotólisis también libera 2H+, a los que se les sumarán los 2 H+ bombeados por la citocromo b6-f desde el estroma al espacio tilacoidal como consecuencia del transporte de electrones, generándose una diferencia de potencial electroquímico que hace que los H+ pasen al estroma a través de la ATP-SINTETASA produciéndose la síntesis de ATP.
Tanto el ATP como el NADPH obtenidos en la fase luminosa acíclica se utilizan en la fase oscura o ciclo de calvin. En el caso del ATP para fosforilar el ácido 3-fosfoglicérico dando lugar al 1,3-difosfoglicérico y gracias al NADPH este se reduce a gliceraldehido-3-fosfato. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (pueden quedarse en el estroma para la síntesis de almidón,.. o puede ir al citosol para dar lugar a la glucosa)

7.- ¿Qué es el ATP? ¿Qué misión fundamental cumple en los organismos? ¿En qué se parece(químicamente a los ácidos nucleicos? ¿Cómo lo sintetizan las células (indicar dos procesos).
El ATP es una coenzima de transferencia, las cuales se encargan de transportar grupos funcionales, en el caso del ATP grupos fosfato. Este actúa como moneda energética y actúa en todas las reacciones de intercambio de energía. Además está compuesto por adenina, tres fosfatos y ribosa (en esto es en lo que se parecen a los ácidos nucleicos, en la composición). Por lo que respecta a los enlaces de esos grupos fosfatos estos requieren de mucha energía, por lo que al sintetizar ATP a partir de ADP y Pi serán capaces de almacenar la energía, mientras que al descomponer el ATP en ADP y Pi esta energía podrá ser liberada.
De esta manera en las reacciones endergónicas (que son en las que se consume energía), lo que va a producirse es la hidrólisis del ATP dando lugar a ADP y Pi, de manera que debido a la ruptura de ese enlace se produce la obtención de 7 Kcal/mol. Sin embargo hay una ruta alternativa por la cual se puede obtener el doble de energía que consiste en la ruptura de los dos enlaces de alta energía del ATP, de manera que este se escinde en AMP y Pirofosfato, y posteriormente ese pirofosfato que son los dos fosfatos inorgánicos juntos se hidroliza. De esta forma podemos obtener el doble de energía es decir 14 Kcal/mol.
Mientras que en las reacciones endergónicas (en las que se genera la energía), se sintetiza ATP a partir de ADP y Pi.
Se puede sintetizar de varias formas 2 de ellas son:
FOTOFOSFORILACIÓN: es la síntesis de ATP que se produce en la membrana de los tilacoides. Lo que ocurre es que al incidir 2 fotones sobre el fotosistema ya sea I o II se liberan 2 electrones, estos pasarán por diferentes aceptores, dando lugar a la cadena transportadora de electrones. Por otro lado los electrones en el caso del fotosistema II perdidos se recuperan por la fotólisis de una molécula de agua. De esa manera lo que ocurre es que el transporte de electrones provoca el bombeo de protones por parte del citocromo B6-F del estroma al interior de los tilacoides, estos se juntan con los de la fotólisis de agua en caso de que haya fotosistemas II y generan una diferencia de potencial electroquímico entre las membranas, que hace que los protones salgan a través de la ATP-SINTETASA, produciéndose la síntesis de ATP.
FOSFORILACIÓN A NIVEL DE SUSTRATO: es la síntesis de ATP gracias a la energía que se libera en la degradación de biomoléculas, mediante la hidrólisis de algunos de sus enlaces ricos en energía. Este proceso es regulado por las QUINASAS.
8.- De los siguientes grupos de organismos, ¿Cuáles llevan a cabo la respiración celular? ¿Cuáles realizan la fotosíntesis oxigénica?: algas eucariotas, angiospermas, cianobacterias (cianofíceas), helechos y hongos.
Respiración celular: angiospermas, algas eucariotas, cianobacterias, helechos y hongos.
Fotosíntesis oxigénica: algas eucariotas, cianobacterias, helechos y angiospermas.
9.- Del orden de un 50 % de la fotosíntesis que se produce en el planeta es debida a la actividad de microorganismos. Indique en qué consiste el proceso de la fotosíntesis. ¿Cuáles son los sustratos necesarios y los productos finales resultantes?
La fotosíntesis es un proceso anabólico en el que la energía luminosa (captada gracias a los pigmentos fotosintéticos que hay en la membrana de los tilacoides de los cloroplastos) se transforma en energía química, la cual se almacena primeramente en forma de ATP para luego ser utilizada para sintetizar moléculas orgánicas más estables. Más concretamente lo que ocurre es que en el ciclo de calvin se fija el CO2 en sustancias orgánicas utilizando el ATP y el NADPH obtenidos en la fase luminosa, para obtener así una molécula orgánica más compleja.
En primer lugar se produce la fase luminosa, en la cual se capta la energía luminosa gracias a los pigmentos fotosintéticos de la membrana de los tilacoides de los cloroplastos. En esta fase se obtienen el ATP y el NADPH que serán utilizados en la siguiente fase, que es la fase oscura, para la síntesis de materia orgánica.
En la fase luminosa acíclica lo que va a ocurrir es que sobre el fotosistema II (que es el primero que interviene) van a incidir 2 fotones que van a excitar su pigmento diana que es la clorofila P680 del centro de reacción. Esta va a perder 2 electrones que recuperará gracias a la hidrólisis de la molécula de agua, que se llama fotólisis del agua, es decir la ruptura de los enlaces del agua, en la que se obtendrá 2H+, 2 electrones y 1/2 de O2. Esos 2 electrones perdidos son captados por el aceptor primario que es la feofitina, luego pasan a la PLASTOQUINONA, esta los cede al citocromo b6-f (que bombea protones del estroma al espacio tilacoidal) y de este pasan a la PLASTOCIANINA. Finalmente son captados por la clorofila P700 del fotosistema I. De esta manera los 2 electrones que la clorofila P700 del fotosistema I ha perdido al incidir sobre el 2 fotones son recuperados. Los 2 electrones perdidos por el fotosistema I pasan a la FERREDOXINA, que los transporta hasta la NADP+ REDUCTASA, donde van a llegar 2H+ del estroma para formar hidrógeno que será captado por el NADP+ para reducirse a NADPH+ (fotorreducción del NADP+).
Mientras que con los H+ lo que va a ocurrir es lo siguiente. La energía desprendida en el transporte de electrones va bombear H+ del estroma al interior del tilacoide (por el citocromo b6-f) y estos junto con los 2 de la fotólisis del agua van a generar una diferencia de potencial electroquímico a ambos lados de la membrana que va a hacer que los protones salgan al estroma a través de la ATP-SINTETASA, produciéndose la fotofosforilación, es decir la formación de ATP.
De esta manera hemos obtenido a partir de una molécula de agua y 4 fotones, 1 NADPH, 1/2 O2 y un ATP. (Por cada 2 electrones entran 4 H+, 2 de la hidrólisis del agua y 2 impulsados por la cadena de transporte,
y tres protones dan lugar a 1 ATP). Sin embargo se requiere de 3 ATP para que se produzca la fase oscura por lo que se deberá llevar a cabo una fase luminosa cíclica.
En la fase luminosa cíclica solo actúa el fotosistema I, cuya clorofila P700 pierde 2 electrones por la incidencia de 2 fotones que volverán otra vez a el, por lo que no hay ni fotólisis del agua, reducción del NADP+, ni se desprende O2. En este caso inciden 2 fotones sobre el fotosistema I que excitan a la clorofila P700, de manera que pierde 2 electrones, estos pasan a la FERREDOXINA, después al CITOCROMO B6-F, (que bombea los protones desde el estroma al interior de los tilacoides, y cuando estos salen por la ATP-SINTETASA, provocan la síntesis de ATP), posteriormente pasan a la PLASTOCIANINA y luego al PSI otra vez. De manera que así hemos obtenido la cantidad de energía que necesitábamos para realizar la fase oscura.

Por tanto el balance de esta fase luminosa es el siguiente: En primer lugar, en la fase acíclica
a partir de una molécula de agua y 4 fotones, obtenemos 1 NADPH, 1/2 O2 y un ATP. (Por cada 2 electrones entran 4 H+, 2 de la hidrólisis del agua y 2 impulsados por la cadena de transporte,
y tres protones dan lugar a 1 ATP). Mientras que en la fase luminosa cíclica solo se produce ATP, pero es muy importante ya que en la fase oscura se requiere de más ATP del que se puede producir en la fase luminosa acíclica. Ese ATP y NADPH obtenidos se utilizan en la fase oscura.
En la fase oscura de la fotosíntesis en la que no se requiere de luz, lo que se produce es la síntesis de materia orgánica a partir de CO2, utilizándose el ATP y el NADPH obtenido en la fase luminosa. Esta fase se da lugar en el estroma de los cloroplastos. A grandes rasgos se podría decir que es la fijación del carbono a partir de CO2, para dar lugar a una molécula orgánica más estable. Este se divide en tres fases que son la fijación del CO2, la reducción del CO2 fijado y la regeneración de la ribulosa 1,5-difosfato, las cuales se llevan a cabo de la siguiente manera:
Primeramente la pentosa ribulosa 1,5-difosfato se une en el estroma al CO2 atmosférico gracias a la enzima ribulosa 1,5-difosfato carboxilasa oxidasa, conocida con el nombre de ribusco, que es la que posibilita esa fijación del CO2 en la pentosa. De esa manera se da lugar a un compuesto inestable de 6 átomos de carbono el cual se divide en 2 moléculas de ácido 3-fosfoglicérico, que son moléculas de 3 átomos de carbono.
Posteriormente, se produce la reducción de ese CO2 fijado. En este caso, primero el ácido 3-fosfoglicérico es fosforilado, consumiéndose de esa manera ATP (obtenido en la fase luminosa, que se reduce a ADP y Pi) y así obtenemos 1,3-difosfoglicérico. Y a continuación este se reduce consumiéndose el NADPH (obtenido en la fase luminosa que se reduce a NADP+) y así obtenemos gliceraldehido 3-fosfato. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (puede quedarse en el estroma para la síntesis de almidón, ácidos grasos, aminoácidos,.. o puede ir al citosol para dar lugar a la glucosa o fructosa).
En el caso de la regeneración de la ribulosa 1,5-difosfato, lo que ocurre es que el gliceraldehido 3-fosfato se transforma en ribulosa-5-fosfato mediante un proceso complejo en el que se suceden una serie de compuestos de 3,4,5 y 7 carbonos. Al final se regenera la ribulosa 1,5-difosfato mediante la fosforilación directa con ATP. (este proceso es conocido como ciclo de las pentosas fosfato)
Por tanto en el ciclo de calvin por cada CO2 que se incorpora se requiere de 2 NADPH y 3 ATP. Este es un proceso que no requiere de energía luminosa ni de clorofila, pero se suele llevar a cabo durante el día porque es cuando se sintetizan el ATP y el NADPH.

10.- Describe la fase luminosa de la fotosíntesis y cuál es su aporte al proceso fotosintético global.
La fase luminosa es un proceso en el que se capta la energía luminosa gracias a los pigmentos fotosintéticos de la membrana de los tilacoides de los cloroplastos. En esta fase se obtienen el ATP y el NADPH que serán utilizados en la siguiente fase, que es la fase oscura, para la síntesis de materia orgánica.
En la fase luminosa acíclica lo que va a ocurrir es que sobre el fotosistema II (que es el primero que interviene) van a incidir 2 fotones que van a excitar su pigmento diana que es la clorofila P680 del centro de reacción. Esta va a perder 2 electrones que recuperará gracias a la hidrólisis de la molécula de agua, que se llama fotólisis del agua, es decir la ruptura de los enlaces del agua, en la que se obtendrá 2H+, 2 electrones y 1/2 de O2. Esos 2 electrones perdidos son captados por el aceptor primario que es la feofitina, luego pasan a la PLASTOQUINONA, esta los cede al citocromo b6-f (que bombea protones del estroma al espacio tilacoidal) y de este pasan a la PLASTOCIANINA. Finalmente son captados por la clorofila P700 del fotosistema I. De esta manera los 2 electrones que la clorofila P700 del fotosistema I ha perdido al incidir sobre el 2 fotones son recuperados. Los 2 electrones perdidos por el fotosistema I pasan a la FERREDOXINA, que los transporta hasta la NADP+ REDUCTASA, donde van a llegar 2H+ del estroma para formar hidrógeno que será captado por el NADP+ para reducirse a NADPH+ (fotorreducción del NADP+).
Mientras que con los H+ lo que va a ocurrir es lo siguiente. La energía desprendida en el transporte de electrones va bombear H+ del estroma al interior del tilacoide (por el citocromo b6-f) y estos junto con los 2 de la fotólisis del agua van a generar una diferencia de potencial electroquímico a ambos lados de la membrana que va a hacer que los protones salgan al estroma a través de la ATP-SINTETASA, produciéndose la fotofosforilación, es decir la formación de ATP.
De esta manera hemos obtenido a partir de una molécula de agua y 4 fotones, 1 NADPH, 1/2 O2 y un ATP. (Por cada 2 electrones entran 4 H+, 2 de la hidrólisis del agua y 2 impulsados por la cadena de transporte,
y tres protones dan lugar a 1 ATP). Sin embargo se requiere de 3 ATP para que se produzca la fase oscura por lo que se deberá llevar a cabo una fase luminosa cíclica.
En esta solo actúa el fotosistema I, cuya clorofila P700 pierde 2 electrones por la incidencia de 2 fotones que volverán otra vez a el, por lo que no hay ni fotólisis del agua, reducción del NADP+, ni se desprende O2. En este caso inciden 2 fotones sobre el fotosistema I que excitan a la clorofila P700, de manera que pierde 2 electrones, estos pasan a la FERREDOXINA, después al CITOCROMO B6-F, (que bombea los protones desde el estroma al interior de los tilacoides, y cuando estos salen por la ATP-SINTETASA, provocan la síntesis de ATP), posteriormente pasan a la PLASTOCIANINA y luego al PSI otra vez. De manera que así hemos obtenido la cantidad de energía que necesitábamos para realizar la fase oscura.
Por tanto el balance de esta fase luminosa es el siguiente: En primer lugar, en la fase acíclica
a partir de una molécula de agua y 4 fotones, obtenemos 1 NADPH, 1/2 O2 y un ATP. (Por cada 2 electrones entran 4 H+, 2 de la hidrólisis del agua y 2 impulsados por la cadena de transporte,
y tres protones dan lugar a 1 ATP). Mientras que en la fase luminosa cíclica solo se produce ATP, pero es muy importante ya que en la fase oscura se requiere de más ATP del que se puede producir en la fase luminosa acíclica. Ese ATP y NADPH obtenidos se utilizan en la fase oscura, el ATP para fosforilar el ácido 3-fosfoglicérico dando lugar al 1,3-difosfoglicérico y gracias al NADPH este se reduce a gliceraldehido-3-fosfato,. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (puede quedarse en el estroma para la síntesis de almidón,.. o puede ir al citosol para dar lugar a la glucosa).
11.- ¿Qué es un organismo autótrofo quimiosintético?
Un organismo autótrofo quimiosintético generalmente es una bacteria que aprovecha la energía desprendida en la oxidación de ciertas moléculas.
12.- Define en no más de cinco líneas el concepto de "Metabolismo", indicando su función biológica.
METABOLISMO: es el conjunto de reacciones químicas que tienen lugar en el interior de las células para satisfacer las necesidades de materia y energía de la misma. Todas estas reacciones se relacionan en unas rutas o vías metabólicas, en las que en cada reacción interviene como mínimo una enzima, de manera que el producto de una reacción es el sustrato de la siguiente. Hay 2 tipos de rutas el anabolismo y el catabolismo. Y el objetivo del metabolismo es la obtención de energía, a través de la degradación de materia orgánica, la obtención de materia precursora sencilla necesaria para la síntesis de materia orgánica y por último la biosíntesis de moléculas orgánicas.
13.- Indique qué frases son ciertas y cuáles son falsas. Justifique la respuesta:
a) Una célula eucariótica fotoautótrofa tiene cloroplastos pero no tiene mitocondrias: Falso, es eucariota por lo tanto también tiene mitocondrias.
b) Una célula eucariótica quimioheterótrofa posee mitocondrias pero no cloroplastos. Verdadero, ya que se trata de células animales, por tanto no tiene cloroplastos.
c) Una célula procariótica quimioautótrofa no posee mitocondrias ni cloroplastos: Verdadero, posee otro tipo de orgánulos.
d) Las células de las raíces de los vegetales son quimioautótrofas: Verdadero, ya que su fuente de energía es la energía que se desprende en las reacciones químicas.
14.- Fotosistemas: Conceptos de complejo antena y centro de reacción. Función y localización.
Los fotosistemas son complejos formados por proteínas transmembranosas que contienen pigmentos fotosintéticos y forman 2 subunidades que son las siguientes:
Complejo captador de luz o antena: en el predominan los pigmentos sobre las proteínas. Estos pigmentos antena (carotenoides, clorofila a y b) solo van a poder captar la energía luminosa y la van a transportar a otros pigmentos del centro de reacción. Este se encuentra en la periferia del fotosistema.
Centro de reacción: en el predominan las proteínas sobre los pigmentos y se encuentra en el interior del fotosistema. En este centro de reacción se encuentra el pigmento diana que son dos moléculas especiales de clorofila a. Es el lugar donde va a parar la energía luminosa captada por los pigmentos antenas. Más concretamente un electrón del pigmento diana va a captar esa energía y va a pasar a un aceptor de electrones, de manera que los fotosistemas deberán recuperar los electrones perdidos si quieren seguir funcionando.
Hay 2 tipos de fotosistemas el I y el II, el I actúa en las dos fases luminosas (cíclica y acíclica) y el II solo en la acíclica. Por otro lado, el I se encuentra casi exclusivamente en los tilacoides de estroma mientras que el dos es más abundante en los tilacoides apilados que forman los grana. Además en el I en su centro de reacción hay dos moléculas de clorofila a que se llaman P700 y en el II se llaman P680.

15.- Compara: a) quimiosíntesis y fotosíntesis b) fosforilación oxidativa y fotofosforilación.
a) Tanto la quimiosíntesis como la fotosíntesis son procesos anabólicos en los que se sintetiza materia orgánica compleja a partir de materia orgánica sencilla, por tanto la molécula inicial de ambos es una molécula orgánica. Sin embaro, el modo en como obtienen la energía es distinto, en la quimiosíntesis la energía se obtiene de la que es desprendida en la oxidación de ciertas sustancias, mientras que en la fotosíntesis se transforma la energía luminosa en energía química. Esto explica porque en la fotosíntesis hay fase luminosa (para la transformación de esa energía) y en la quimiosíntesis no, pero en ambas se produce el ciclo de calvin, es decir en ambas se fija el CO2 a la molécula orgánica para dar lugar a una molécula más estable.
b) En el caso de la fosforilación oxidativa y la fotofosforilación se trata de procesos muy similares con unas diferencias puntuales. Ambos son un tipo de síntesis de ATP que se produce gracias a la energía que se libera en el transporte de electrones a lo largo de la cadena de transporte electrónico, este transporte lo que hace es generar un bombeo de protones que a su vez genera un gradiente electroquímico que hace que los protones sean bombeados otra vez al lugar de donde habían salido bombeados anteriormente, este flujo se produce a través de la ATP-SINTETASA y da lugar a la síntesis de ATP. Sin embargo en ese proceso que es comparten ambas fosforilaciones encontramos ciertas diferencias. En primer lugar, la cadena de transporte electrónico en la fotofosforilación se encuentra en la membrana de los tilacoides de los cloroplastos, mientras que en la fosforilación oxidativa se encuentra en la membrana mitocondrial interna más concretamente en las crestas mitocondriales. Por otro lado, en la fosforilación oxidativa los electrones son cedidos por el NADH y FADH2 obtenidos en las reacciones anteriores, mientras que en la fotofosforilación los electrones son perdidos por los fotosistemas al incidir sobre ellos fotones de luz. Además en la fosforilación oxidativa cuando los protones son bombeados por primera vez pasan de la matriz mitocondrial al espacio intermembrana y después por el gradiente electroquímico vuelven a entras en la matriz mitocondrial ,mientras que en la fotofosforilación primeramente los protones se bombean del estroma al espacio tilacoidal y después por el gradiente electroquímico pasan otra vez al estroma, sintetizándose el ATP.
16.- La vaca utiliza los aminoácidos de la hierba para sintetizar otras cosas, por ejemplo la albúmina de la leche (lactoalbúmina). Indica si este proceso será anabólico o catabólico. Razona la respuesta.
Es un proceso anabólico, más concretamente se trata del anabolismo de proteínas pues en el proceso que se está produciendo a partir de moléculas más sencillas, que en este caso son los aminoácidos, se da lugar a moléculas orgánicas más complejas como son en este caso las proteínas albúminas.
17.- Explica brevemente si la proposición que sigue es verdadera o falsa. El ATP es una molécula dadora de energía y de grupos fosfatos.
Dicha afirmación es verdadera ya que conocemos las siguientes ideas acerca del ATP. El ATP es una coenzima de transferencia, las cuales se encargan de transportar grupos funcionales, en el caso del ATP grupos fosfato. Este actúa como moneda energética y actúa en todas las reacciones de intercambio de energía. Además está compuesto por adenina, tres fosfatos y ribosa (en esto es en lo que se parecen a los ácidos nucleicos, en la composición). Por lo que respecta a los enlaces de esos grupos fosfatos estos requieren de mucha energía, por lo que al sintetizar ATP a partir de ADP y Pi serán capaces de almacenar la energía, mientras que al descomponer el ATP en ADP y Pi esta energía podrá ser liberada.
De esta manera en las reacciones endergónicas (que son en las que se consume energía), lo que va a producirse es la hidrólisis del ATP dando lugar a ADP y Pi, de manera que debido a la ruptura de ese enlace se produce la obtención de 7 Kcal/mol. Sin embargo hay una ruta alternativa por la cual se puede obtener el doble de energía que consiste en la ruptura de los dos enlaces de alta energía del ATP, de manera que este se escinde en AMP y Pirofosfato, y posteriormente ese pirofosfato que son los dos fosfatos inorgánicos juntos se hidroliza. De esta forma podemos obtener el doble de energía es decir 14 Kcal/mol.
Mientras que en las reacciones endergónicas (en las que se genera la energía), se sintetiza ATP a partir de ADP y Pi.

18.- ¿En qué lugar de la célula y de qué manera se puede generar ATP?
Se puede generar en el citosol, en este caso se trata de la fosforilación a nivel de sustrato, que se produce en reacciones como la glucólisis. peo también en el ciclo de krebs, que se lleva a cabo en la matriz mitocondrial. Y su producción se lleva a cabo gracias a la energía que se libera en la degradación de biomoléculas, mediante la hidrólisis de algunos de sus enlaces ricos en energía. Este proceso es regulado por las QUINASAS.
También se puede generar en las cadenas respiratorias mitocondriales (crestas mitocondriales). En este caso se llama fosforilación oxidativa, que es la síntesis de ATP gracias a la energía liberada en el transporte de electrones en la cadena respiratoria de las mitocondrias. Este transporte provoca un bombeo de protones al espacio intermembrana, generándose una diferencia de gradiente electroquímico que impulsa los electrones de regreso a la matriz mitocondrial a través de la ATP-SINTETASA. De manera que la energía liberada por el flujo de H+ a favor de gradiente posibilita la unión de ADP y Pi.
Y por último también se puede generar en las cadenas respiratorias de los fotosistemas I y II, que se encuentran en la membrana de los tilacoides. A esta se le llama fotofosforilación, y consiste en lo siguiente. Es la síntesis de ATP que se produce en la membrana de los tilacoides. Lo que ocurre es que al incidir 2 fotones sobre el fotosistema ya sea I o II se liberan 2 electrones, estos pasarán por diferentes aceptores, dando lugar a la cadena transportadora de electrones. Por otro lado los electrones en el caso del fotosistema II perdidos se recuperan por la fotólisis de una molécula de agua. De esa manera lo que ocurre es que el transporte de electrones provoca el bombeo de protones por parte del citocromo B6-F del estroma al interior de los tilacoides, estos se juntan con los de la fotólisis de agua en caso de que haya fotosistemas II y generan una diferencia de potencial electroquímico entre las membranas, que hace que los protones salgan a través de la ATP-SINTETASA, produciéndose la síntesis de ATP.
19.- Papel del acetil-CoA en el metabolismo. Posibles orígenes del acetil-CoA celular y posibles destinos metabólicos (anabolismo y catabolismo). Principales rutas metabólicas que conecta.
En el metabolismo el Acetil-CoA tiene un gran protagonismo, y es que en este es donde convergen todas las rutas catabólicas (glucólisis, B-oxidación y transaminación oxidativa), además es el punto de partida del ciclo de krebs, ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos. De esa manera tiene un papel fundamental en el catabolismo y en el anabolismo,que es posibilitar la degradación de glúcidos, lípidos, proteínas,...así como también permite la síntesis de moléculas más complejas.
De esta manera, en el caso del catabolismo los orígenes del acetil-CoA pueden ser los siguientes: la degradación de los ácidos grasos,(a través de la hélice de Lynen), el catabolismo de las proteínas y el catabolismo de los azúcares (tras haberse producido la glucólisis y la descarboxilación oxidativa). Por lo que respecta a su destino, es la oxidación total en el ciclo de krebs, dando lugar a CO2, NADH , FADH2 y GTP.
En el caso del anabolismo, el acetil-CoA lo veremos en el anabolismo de los lípidos, este es el precursor de la obtención de los ácidos grasos, primeramente se produce en las mitocondrias tras la descarboxilación del ácido pirúvico y después para salir al citoplasma donde se encuentra el complejo SAG, se unirá a un oxalacetato para dar citrato que sale al citosol, entonces se libera el Acetil-CoA. Este acetil-CoA se transformará en malonil, que es el que permite que se vayan incorporando átomos de carbono.
20.- Esquematiza la glucólisis: a) Indica al menos, sus productos iniciales y finales. b) Destino de los productos finales en condiciones aerobias y anaerobias. c) Localización del proceso en la célula.

21.- Una célula absorbe n moléculas de glucosa y las metaboliza generando 6n moléculas de CO2 y consumiendo O2 .¿ Está la célula respirando ? ¿Para qué? ¿participa la matriz mitocondrial? ¿Y las crestas mitocondriales?.
Si, la célula está respirando ya que el enunciado nos muestra claramente que el proceso que está llevando a cabo la célula es la respiración celular, es decir, la degradación completa de la glucosa en presencia de oxígeno atmosférico, obteniéndose como resultado agua, CO2 y energía en forma de ATP necesaria para diversos procesos celulares (para eso se produce la respiración). Este proceso se da en la matriz mitocondrial, así como también se da en esta la descarboxilación oxidativa. Y por otro lado las crestas mitocondriales, que se encuentran en la membrana mitocondrial interna también actúan ya que es en ellas donde se produce la cadena de transporte electrónico gracias a la cual se libera la energía contenida en los coenzimas NADH y FADH2 obtenidos en las reacciones anteriores, generándose ATP a partir de ADP y Pi.
22.- ¿Qué ruta catabólica se inicia con la condensación del acetil-CoA y el ácido oxalacético, y qué se origina en dicha condensación? ¿De dónde provienen fundamentalmente cada uno de los elementos? ¿Dónde tiene lugar esta ruta metabólica?.
Con la condensación del acetil- CoA (2 átomos de carbono) y el ácido oxalacético (4 átomos de carbono) se inicia el ciclo de krebs, ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos, el cual se lleva a cabo en la matriz mitocondrial, y el resultado de esa condensación es la formación de un ácido tricarboxílico que está formado por 6 átomos de carbono que es el ácido cítrico (de ahí el nombre del ciclo), y también se desprende un Co-A.
En primer lugar, el acetil- CoA puede provenir de varias rutas de distintos tipos de catabolismo. Uno de ellos es el catabolismo de las proteínas, otro es el catabolismo de los ácidos grasos. Y por otro lado siguiendo la ruta del catabolismo de los azúcares también podemos llegar a este acetil-CoA, a través de la descarboxilación oxidativa.
23.- ¿Qué molécula acepta el CO2 en la fotosíntesis? ¿Qué enzima cataliza esta reacción? ¿A qué moléculas da lugar?.
La molécula que acepta el CO2 en la fotosíntesis es la pentosa ribulosa-1,5-difosfato, y la enzima que cataliza esta reacción, es decir la que posibilita la fijación del CO2 es la ribulosa-1,5-difosfato carboxilasa oxidasa, conocida con el nombre de ribusco. De esa manera se da lugar a un compuesto inestable de 6 átomos de carbono el cual se divide en 2 moléculas de ácido 3-fosfoglicérico, que son moléculas de 3 átomos de carbono. Posteriormente se produce la reducción de ese CO2 fijado. Primeramente el ácido 3-fosfoglicérico es fosforilado, consumiéndose de esa manera ATP (obtenido en la fase luminosa, que se reduce a ADP y Pi) y obteniéndose 1,3-difosfoglicérico. Y a continuación este se reduce consumiéndose el NADPH (obtenido en la fase luminosa que se reduce a NADP+) y obteniéndose gliceraldehido 3-fosfato. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (puede quedarse en el estroma para la síntesis de almidón, ácidos grasos, aminoácidos,.. o puede ir al citosol para dar lugar a la glucosa). En el caso de la regeneración de la ribulosa 1,5-difosfato, lo que ocurre es que el gliceraldehido 3-fosfato se transforma en ribulosa-5-fosfato mediante un proceso complejo en el que se suceden una serie de compuestos de 3,4,5 y 7 carbonos. Al final se regenera la ribulosa 1,5-difosfato mediante la fosforilación directa con ATP. (este proceso es conocido como ciclo de las pentosas fosfato)

24.- Indique cuál es el papel biológico del NAD, NADH + H. en el metabolismo celular. Escriba tres reacciones en las cuáles participe.
El NAD es una coenzima de oxidación y reducción, que se encarga de transportar protones y electrones, por lo que su función fundamental es el almacenamiento y transporte de energía. Entre las reacciones en las que participa se encuentran la glucólisis, la descarboxilación oxidativa o el ciclo de krebs.
25.- Explique brevemente el esquema siguiente:
El siguiente esquema muestra el ciclo de calvin, es decir la fase oscura de la fotosíntesis que se produce tras haberse llevado a cabo la fase luminosa ya que requiere del ATP y NADPH que se producen en esta. La molécula que acepta el CO2 en el ciclo de calvin es la pentosa ribulosa-1,5-difosfato, y la enzima que cataliza esta reacción, es decir la que posibilita la fijación del CO2 es la ribulosa-1,5-difosfato carboxilasa oxidasa, conocida con el nombre de ribusco. De esa manera se da lugar a un compuesto inestable de 6 átomos de carbono el cual se divide en 2 moléculas de ácido 3-fosfoglicérico, que son moléculas de 3 átomos de carbono. Posteriormente se produce la reducción de ese CO2 fijado. Primeramente el ácido 3-fosfoglicérico es fosforilado, consumiéndose de esa manera ATP (obtenido en la fase luminosa, que se reduce a ADP y Pi) y obteniéndose 1,3-difosfoglicérico. Y a continuación este se reduce consumiéndose el NADPH (obtenido en la fase luminosa que se reduce a NADP+) y obteniéndose gliceraldehido 3-fosfato. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (puede quedarse en el estroma para la síntesis de almidón, ácidos grasos, aminoácidos, glicerina,... o puede ir al citosol para dar lugar a la glucosa).

26.- Bioenergética: a) Defina los conceptos de: fosforilación a nivel del sustrato, fotofosforilación y fosforilación oxidativa. b) ¿En qué niveles celulares se produce cada uno de dichos mecanismos y por qué?
FOSFORILACIÓN A NIVEL DE SUSTRATO: es la síntesis de ATP gracias a la energía que se libera en la degradación de biomoléculas, mediante la hidrólisis de algunos de sus enlaces ricos en energía. Este proceso es regulado por las QUINASAS.
FOSFORILACIÓN OXIDATIVA: es un proceso similar al anterior en este caso es la síntesis de ATP gracias a la energía liberada en el transporte de electrones en la cadena respiratoria de las mitocondrias. Este transporte provoca un bombeo de protones al espacio intermembrana, generándose una diferencia de gradiente electroquímico que impulsa los electrones de regreso a la matriz mitocondrial a través de la ATP-SINTETASA. De manera que la energía liberada por el flujo de H+ a favor de gradiente posibilita la unión de ADP y Pi. Por tanto en eucariotas se da en la membrana mitocondial interna, mientras que en las células procariotas se da en la membrana plasmática.
FOTOFOSFORILACIÓN: es la síntesis de ATP que se produce en la membrana de los tilacoides de los cloroplastos. Lo que ocurre es que al incidir 2 fotones sobre el fotosistema ya sea I o II se liberan 2 electrones, estos pasarán por diferentes aceptores, dando lugar a la cadena transportadora de electrones. Por otro lado los electrones en el caso del fotosistema II perdidos se recuperan por la fotólisis de una molécula de agua. De esa manera lo que ocurre es que el transporte de electrones provoca el bombeo de protones por parte del citocromo B6-F del estroma al interior de los tilacoides, estos se juntan con los de la fotólisis de agua en caso de que haya fotosistemas II y generan una diferencia de potencial electroquímico entre las membranas, que hace que los protones salgan a través de la ATP-SINTETASA, produciéndose la síntesis de ATP.
27.- Describa el proceso de transporte electrónico mitocondrial y el proceso acoplado de fosforilación oxidativa. Resuma en una reacción general los resultados de ambos procesos acoplados. A la luz de lo anterior, ¿Cuál es la función metabólica de la cadena respiratoria? ¿Por qué existe la cadena respiratoria? ¿Dónde se localiza?.
Tras haberse producido el ciclo de calvin, parte de la energía de la glucosa se encuentra todavía retenida en los electrones de alta energía de los coenzimas reducidos NADH y FADH2 obtenidos en las anteriores etapas. Esta energía será liberada en la cadena de transporte de electrones cuando el NADH y el FADH2 cedan sus electrones a una de las miles de cadenas de transporte electrónico que hay distribuidas por la membrana interna de las mitocondrias, los cuales van pasar a través de los diferentes complejos que componen a la cadena, que son 4 grandes complejos proteicos, una molécula lipídica pequeña (UBIQUINONA) y una molécula proteica pequeña (CITOCROMO C). Esta cadena va a transportar los electrones hasta el OXÍGENO que es el último aceptor ÚLTIMO ACEPTOR de electrones, el cual se reduce para dar lugar a H2O que es uno de los productos finales de la degradación de la glucosa. Ese transporte de electrones se produce mediante una secuencia de reacciones redox, en las que intervienen 2 transportadores uno cede un par de electrones reduciéndose y el otro los capta oxidándose. Esos transportadores a su vez no están colocados de manera irregular sino que se colocan de menor a mayor potencial redox. Por tanto el transporte de electrones se produce a favor de diferencia de POTENCIAL REDOX (proceso fuertemente EXERGÓNICO, en el que se libera una gran cantidad de energía).
El NADH cede sus electrones al complejo I, bombeando 10 H+ al espacio intercelular. Sin embargo el FADH2 los cede a la ubiquinona por lo que eluden el complejo I, y por tanto solo bombean 6 H+. Hay tener en cuenta que el NADH obtenido en el citosol tendrá que pasar al interior de la mitocondria a través de un SISTEMA DE LANZADERA.
La energía que se libera en este transporte es aprovechada por la célula mediante el acoplamiento de una fosforilación oxidativa al transporte electrónico.
El proceso por tanto consiste en que primeramente el transporte de electrones (MOVIMIENTO DE ELECTRONES) provoca el bombeo de protones de la matriz mitocondrial al espacio intermembrana (QUIMIÓSMOSIS) y este bombeo genera un gradiente electroquímico (en el que se almacena la energía liberada en el transporte de electrones). De esa forma los protones van a tender a entrar otra vez en la matriz mitocondrial a tráves de las ATP-SINTETASA que se encuentran en la cadena respiratoria. Esta entrada se va a producir a favor de gradiente electroquímico. Por lo que gracias al flujo de protones a través de la ATP-SINTETASA a favor de gradiente electroquímico se va a liberar la energía que va a fosforilar el ADP y Pi a ATP. (FOSFORILACIÓN OXIDATIVA).
28.- ¿Qué tipos y cuántas moléculas se consumen y se liberan en cada una de las vueltas de la espiral de Lynen en la B-oxidación de los ácidos grasos?.
Cuando los triacilgliceridos o grasas neutras se separan (en el citosol) en glicerina y los 3 ácidos grasos, lo que va a ocurrir con esos ácidos grasos es que van a penetrar en la matriz mitoncondrial donde se va a llevar a cabo la B-oxidación de estos. Para ello primeramente deben ser activados por el coenzima-A transformándose en Acil(graso)-CoA. Esta activación tiene lugar cuando el ácido graso atraviesa la membrana mitocondrial externa y requiere de ATP, de manera que este se hidroliza para dar AMP y pirofosfato, que posteriormente se dividirá en dos Pi, así se han consumido dos enlaces fosfato alto energéticos. Por lo que aquí ya tenemos el consumo de una molécula de ATP y un Coenzima-A para activar al ácido graso. (solo se activa una vez, es decir que cuando se produce la héliza de Lynen no tiene que volver a activarse conforme vaya completando la vuelta porque ya lo está)
Una vez activado ya se produce la vuelta a la hélice de Lynen, en esta primeramente aparece un doble enlace entre el carbono alfa y beta, en este proceso el FADH se reduce a FADH2 (una de las moléculas que se libera). Posteriormente y mediante la incorporación de una molécula de H2O (otra de las moléculas que se consume), el OH de esta se une al carbono beta. Y a continuación ese OH se transforma en un grupo cetona, en este proceso se produce la reducción del NAD a NADH (otra de las moléculas que se libera). Y por último el acil-CoA sufre la pérdida del Acetil-CoA (otra de las moléculas que se libera). En ese momento obtenemos un ácido graso con 2 átomos de carbono menos y este continúa realizando tantas vueltas como sea necesario hasta que la oxidación del ácido graso sea completa. De manera que si se trata de un ácido graso de 14 átomos de carbono se obtendrán 14 acetil- CoA y se darán 13 vueltas, por lo que se obtendrán 13 FADH2 y 13 NADH.
29.- ¿Cómo se origina el gradiente electroquímico de protones en la membrana mitocondrial interna?
Se origina por el bombeo de protones de la matriz mitocondrial al espacio intermembrana, que a su vez es generado por el transporte de electrones en la cadena transportadora de electrones. Estos electrones son cedidos por los coenzimas reducidos NADH y FADH2 obtenidos en las reacciones anteriores a la cadena respiratoria mitondrial, de manera que estos van pasar a través de los diferentes complejos que componen a la cadena, que son 4 grandes complejos proteicos, una molécula lipídica pequeña (ubiquinona) y una molécula proteica pequeña (citocromo c). En el caso de los electrones del NADH, estos son cedidos al complejo I (por lo que por cada NADH se bombean 10 H+ al espacio intermembrana) y en el caso de los del FADH2 estos son cedidos a la ubiquinona, que esta a continuación del complejo I (por lo que por cada FADH2 se bombean 6 H+ al espacio intermembrana, ya que se elude el primer centro de bombeo que es el complejo I). Estos electrones irán pasando por la cadena de transporte y en ese transporte se va a desprender energía. Esta energía las células la aprovechan a través del acoplamiento de una fosforilación a ese transporte de electrones. El proceso por tanto consiste en que primeramente el transporte de electrones provoca el bombeo de protones de la matriz mitocondrial al espacio intermembrana y este bombeo genera un gradiente electroquímico (en el que se almacena la energía liberada en el transporte de electrones). De esa forma los protones van a tender a entrar otra vez en la matriz mitocondrial a tráves de las ATP-SINTETASA que se encuentran en la cadena respiratoria. Esta entrada se va a producir a favor de gradiente electroquímico. Por lo que gracias al flujo de protones a través de la ATP-SINTETASA a favor de gradiente electroquímico se va a liberar la energía que va a fosforilar el ADP y Pi a ATP.

30.- ¿Cuál es la primera molécula común en las rutas catabólicas de los glúcidos y los lípidos? ¿Cuál es el destino final de dicha molécula en el metabolismo?
La primera molécula común en las rutas metabólicas de los glúcidos y los lípidos es el Acetil-CoA, cuyo destino es su descomposición total en el ciclo de krebs, ciclo del ácido cítrico o ciclo de los ácidos tricarboxíllicos (para dar lugar a NADH, FADH2, CO2 Y GTP), el cual se inicia con la condensación del acetil-CoA y el ácido oxalacético para dar lugar a un compuesto formado por 6 átomos de carbono que es el ácido cítrico. El NADH y el FADH2 son cedidos a la cadena de transporte de electrones para la obtención de energía.
31.- Ciclo de Calvin: concepto, fases y rendimiento neto.
Es la fase oscura de la fotosíntesis en la que no se requiere de luz, en ella lo que ocurre es la síntesis de materia orgánica a partir de CO2, utilizándose el ATP y el NADPH obtenido en la fase luminosa. Esta fase se da lugar en el estroma de los cloroplastos. A grandes rasgos se podría decir que es la fijación del carbono a partir de CO2, para dar lugar a una molécula orgánica más estable. Este se divide en tres fases que son la fijación del CO2, la reducción del CO2 fijado y la regeneración de la ribulosa 1,5-difosfato, las cuales se llevan a cabo de la siguiente manera:
Primeramente la pentosa ribulosa 1,5-difosfato se une en el estroma al CO2 atmosférico (FIJACIÓN DEL CO2) gracias a la enzima ribulosa 1,5-difosfato carboxilasa oxidasa, conocida con el nombre de ribusco, que es la que posibilita esa fijación del CO2 en la pentosa. De esa manera se da lugar a un compuesto inestable de 6 átomos de carbono el cual se divide en 2 moléculas de ácido 3-fosfoglicérico, que son moléculas de 3 átomos de carbono.
Posteriormente, se produce la REDUCCIÓN DEL CO2 FIJADO. En este caso, primero el ácido 3-fosfoglicérico es fosforilado, consumiéndose de esa manera ATP (obtenido en la fase luminosa, que se reduce a ADP y Pi) y así obtenemos 1,3-difosfoglicérico. Y a continuación este se reduce consumiéndose el NADPH (obtenido en la fase luminosa que se reduce a NADP+) y así obtenemos gliceraldehido 3-fosfato. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (puede quedarse en el estroma para la síntesis de almidón, ácidos grasos, aminoácidos,.. o puede ir al citosol para dar lugar a la glucosa o fructosa).
En el caso de la REGENERACIÓN DE LA RIBULOSA 1,5-DIFOSFATO, lo que ocurre es que el gliceraldehido 3-fosfato se transforma en ribulosa-5-fosfato mediante un proceso complejo en el que se suceden una serie de compuestos de 3,4,5 y 7 carbonos. Al final se regenera la ribulosa 1,5-difosfato mediante la fosforilación directa con ATP. (este proceso es conocido como ciclo de las pentosas fosfato)
Por tanto en el ciclo de calvin por cada CO2 que se incorpora se requiere de 2 NADPH y 3 ATP. Este es un proceso que no requiere de energía luminosa ni de clorofila, pero se suele llevar a cabo durante el día porque es cuando se sintetizan el ATP y el NADPH.
32.- Existe una clase de moléculas biológicas denominadas ATP, NAD, NADP: a) ¿Qué tipo de moléculas son ? (Cita el grupo de moléculas al que pertenecen) ¿Forman parte de la estructura del ADN o del ARN?. b) ¿Qué relación mantienen con el metabolismo celular? (Explícalo brevemente).
a) El ATP, NAD, NADP, son coenzimas, en el caso del NAD y el NADP son coenzimas de oxido reducción que transportan protones y electrones, participando en procesos como la respiración celular. Y en el caso del ATP es una coenzima de transferencia, la cual transporta grupos funcionales, esta acumula energía en sus enlaces y es capaz de cederla para la síntesis de moléculas. Forman la parte no proteica de las enzimas, que se llama cofactor y no forman parte del ADN ni del ARN.
b) Dentro del metabolismo, actúan en reacciones de oxido reducción, lo que hacen es almacenar y transportar energía. En el caso del ATP almacena energía y por lo que respecta al NAD y al NADP, estos transportan electrones (pueden captar electrones, reduciéndose, o pueden ceder electrones, oxidándose).
34.- Balance energético de la degradación completa de una molécula de glucosa.
En primer lugar en la glucolisis, que se produce en el citosol, obtenemos 2 NADH y 2 ATP, realmente se obtienen 4ATP pero en la primera etapa de la glucólisis se consumen 2 ATP. A continuación en la descarboxilación oxidativa, que ya corresponde a la respiración celular y se lleva a cabo en la matriz mitocondrial, se obtienen 2 NADH, que se deberán multiplicar por 2 ya que en la glucólisis obtenemos 2 ácidos pirúvicos, así realmente obtenemos 4 NADH. Posteriormente en el ciclo de krebs obtenemos (por vuelta) 1GTP (análogo al ATP), 3 NADH y un FADH2, todos ellos también se deben multiplicar por 2 ya que en la descarboxilación oxidativa obtenemos 2 moléculas de acetil-CoA, así realmente obtenemos 2 GTP, 6 NADH y 2 FADH2. De manera que en la cadena respiratoria que se producirá posteriormente, por cada NADH se forman 3 ATP y por cada FADH2 se forman 2 ATP, así si tenemos un total de 4 ATP, 10 NADH y 2 FADH2 obtendremos 38 ATP. Sin embargo, en eucariotas se obtienen 36 ATP ya que aunque el paso del ácido pirúvico de la glucólisis a través de permeasas específicas de la membrana mitocondrial no supone un gasto de energía, el NADH que se obtiene en esta glucólisis no puede atravesar la membrana por lo que tiene que ceder sus electrones a un sistema de lanzadera que los cederá a un FADH2 en la matriz mitocondrial para que los lleve a la cadena transportadora de electrones, por lo que se produce la pérdida de 2 ATP.

35.- La siguiente molécula representa el acetil CoA: H3 C-CO-S-CoA.
a) ¿En qué rutas metabólicas se origina y en cuáles se utiliza esta molécula?
De esta manera, en el caso del catabolismo los orígenes del acetil-CoA pueden ser los siguientes: la degradación de los ácidos grasos,(a través de la hélice de Lynen), el catabolismo de las proteínas y el catabolismo de los azúcares (tras haberse producido la glucólisis y la descarboxilación oxidativa). Por lo que respecta a su destino, es la oxidación total en el ciclo de krebs, dando lugar a CO2, NADH , FADH2 y GTP.
En el caso del anabolismo, el acetil-CoA lo veremos en el anabolismo de los lípidos, este es el precursor de la obtención de los ácidos grasos, primeramente se produce en las mitocondrias tras la descarboxilación del ácido pirúvico y después para salir al citoplasma donde se encuentra el complejo SAG, se unirá a un oxalacetato para dar citrato que sale al citosol, entonces se libera el Acetil-CoA. Este acetil-CoA se transformará en malonil, que es el que permite que se vayan incorporando átomos de carbono.
b) De los siguientes procesos metabólicos: Glucogénesis, fosforilación oxidativa y Boxidación, indica: - Los productos finales e iniciales. - Su ubicación intracelular
Glucogénesis: producto inicial (ácido pirúvico, aminoácidos, glicerina o ácido láctico) y producto final (glucosa). Se da en la matriz mitocondrial pero también en el citosol.
Fosforilación oxidativa: producto inicial ADP y Pi, producto final ATP. Se da en la cadena de transporte de electrones, que en la célula animal esta en las crestas mitocondriales de la membrana interna de las mitocondrias, y en las células vegetales se da en la membrana de las tilacoides de los cloroplastos.
B-oxidación de ácidos grasos: producto inicial (ácidos grasos) y producto final (acetil-CoA). Se da en la matriz mitocondrial.
c) Explica con un esquema cómo se puede transformar un azúcar en una grasa ¿Pueden los animales realizar el proceso inverso?
No, ya que los animales carecen de los enzimas que se requieren para transformar una grasa en un azúcar.

36.- En el siguiente diagrama se esquematiza el interior celular y algunas transformaciones de moléculas que se producen en diferentes rutas metabólicas:
a) ¿Qué es el metabolismo? ¿Qué entiendes por anabolismo y catabolismo? ¿Cómo se relacionan el anabolismo y el catabolismo en el funcionamiento de las células? ¿Qué rutas distingues? (Cita sus nombres e indica, si existen, cuáles son los productos inicial y final de cada una de ellas).
El metabolismo es el conjunto de reacciones químicas que tienen lugar en el interior de las células para satisfacer las necesidades de materia y energía de la misma. Todas estas reacciones se relacionan en unas rutas o vías metabólicas, en las que en cada reacción interviene como mínimo una enzima, de manera que el producto de una reacción es el sustrato de la siguiente. Hay 2 tipos de rutas el anabolismo y el catabolismo. Y el objetivo del metabolismo es la obtención de energía, a través de la degradación de materia orgánica, la obtención de materia precursora sencilla necesaria para la síntesis de materia orgánica y por último la biosíntesis de moléculas orgánicas.
Por un lado, el catabolismo es un conjunto de reacciones del metabolismo cuyo objetivo es la obtención de moléculas precursoras sencillas y energía, a partir de la degradación de biomoléculas.
Por otro lado, el anabolismo es una vía constructiva del metabolismo, es decir, es la ruta en la que se produce la síntesis de moléculas complejas a partir de moléculas sencillas y energía.
Ambas rutas están relacionadas ya que la energía y las moléculas precursoras obtenidas en el catabolismo son necesarias para el desarrollo de las reacciones anabólicas.
Una de las rutas que se pueden identificar en la imagen es la glucólisis, en la que el producto inicial es la glucosa y el final es el ácido pirúvico. Otra de las rutas es la descarboxilación oxidativa, en la que el producto inicial es el ácido pirúvico y el NAD+, y el producto final es el acetil-CoA y el NADH. Además también podemos ver el ciclo de krebs en el que el producto inicial es el ácido oxalacético y el acetil-CoA, y los productos finales son 6 NADH, 2 FADH2, 2 GTP y 4 CO2 (cada 2 vueltas al ciclo).
Por último también podemos observar una fermentación láctica, en la que los productos iniciales son ácido pirúvico y NADH y los productos finales son ácido láctico y NAD+.
b) ¿Qué compartimentos celulares intervienen en el conjunto de las reacciones? (Indica el nombre de los compartimentos y la reacción que se produce en cada uno de ellos).
Los 2 compartimentos de la célula que intervienen son las mitocondrias (donde se da la descarboxilación oxidativa y el ciclo de krebs. Y por otro lado el citosol que es donde se da la glucólisis y la fermentación láctica.

37.- Indique el rendimiento energético de la oxidación completa de la glucosa y compárelo con el obtenido en su fermentación anaerobia. Explique las razones de esta diferencia.
En primer lugar en la glucolisis, que se produce en el citosol, obtenemos 2 NADH y 2 ATP, realmente se obtienen 4ATP pero en la primera etapa de la glucólisis se consumen 2 ATP. A continuación en la descarboxilación oxidativa, que ya corresponde a la respiración celular y se lleva a cabo en la matriz mitocondrial, se obtienen 2 NADH, que se deberán multiplicar por 2 ya que en la glucólisis obtenemos 2 ácidos pirúvicos, así realmente obtenemos 4 NADH. Posteriormente en el ciclo de krebs obtenemos (por vuelta) 1GTP (análogo al ATP), 3 NADH y un FADH2, todos ellos también se deben multiplicar por 2 ya que en la descarboxilación oxidativa obtenemos 2 moléculas de acetil-CoA, así realmente obtenemos 2 GTP, 6 NADH y 2 FADH2. De manera que en la cadena respiratoria que se producirá posteriormente, por cada NADH se forman 3 ATP y por cada FADH2 se forman 2 ATP, así si tenemos un total de 4 ATP, 10 NADH y 2 FADH2 obtendremos 38 ATP. Sin embargo, en eucariotas se obtienen 36 ATP ya que aunque el paso del ácido pirúvico de la glucólisis a través de permeasas específicas de la membrana mitocondrial no supone un gasto de energía, el NADH que se obtiene en esta glucólisis no puede atravesar la membrana por lo que tiene que ceder sus electrones a un sistema de lanzadera que los cederá a un FADH2 en la matriz mitocondrial para que los lleve a la cadena transportadora de electrones, por lo que se produce la pérdida de 2 ATP. Sin embargo, en el caso de las fermentaciones solo se obtienen los 2 ATP de la glucólisis, esto se debe a que en las fermentaciones no hay cadena transportadora de electrones, de manera que la fosforilación que se produce siempre es a nivel de sustrato, ya que tampoco hay ATP-SINTETASA (que se encuentra en la cadena transportadora de electrones, esto explica el escaso rendimiento energéticos de las fermentaciones.
38.- ¿En qué orgánulos celulares tiene lugar la cadena de transporte de electrones , uno de cuyos componentes son los citocromos? ¿Cuál es el papel del oxígeno en dicha cadena? ¿Qué seres vivos y para qué la realizan?
Los orgánulos en los que tiene lugar la cadena de transporte electrónico son la mitocondria (membrana mitocondrial interna, crestas mitocondriales) y los cloroplastos (membrana de los tilacoides, en células vegetales), en el caso de las células eucariotas. En el caso de las células procariotas se produce en la membrana plasmática. El oxígeno en esta cadena de transporte actúa como último aceptor de electrones, reduciéndose para formar H2O, que es uno de los productos finales de la degradación de la glucosa.
La llevan a cabo todos los seres vivos aerobios para la obtención de energía.
39.- En el ciclo de Krebs o de los ácidos tricarboxílicos: -¿Qué tipos principales de reacciones ocurren?. - ¿Qué rutas siguen los productos liberados?.
El acetil-CoA obtenido puede oxidarse completamente en la misma mitocondria por el CICLO DE KREBS, CICLO DEL ÁCIDO CÍTRICO O CICLO DE LOS ÁCIDOS TRICARBOXÍLICOS. Este se inicia con la condensación del acetil-CoA (2C), con el ácido oxalacético (4C), dando lugar a un compuesto formado por 6C que es el ácido cítrico. En este proceso también se libera un coenzima A. A continuación se produce una secuencia de 7 reacciones consecutivas en las que se desprende carbono en forma de CO2 y se regenera el ácido oxalacético.
Primero el ácido cítrico pasa a su isómero isocítrico desprendiéndose una molécula de H2O. Posteriormente el ISOCÍTRICO se transforma en ÁCIDO ALFA-CETOGLUTÁRICO (aparece una cetona), reduciéndose el NAD+ a NADH, y desprendiéndose carbono en forma de CO2, es decir se produce una DESCARBOXILACIÓN OXIDATIVA. A continuación, este compuesto vuelve a sufrir una descarboxilación, reduciéndose el NAD+ a NADH y desprendiéndose CO2, así se obtiene el SUCCINIL-CoA. Este pasa en una fosforilación a nivel de sustrato a ÁCIDO SUCCINICO, formándose GTP, análogo al ATP. Posteriormente este compuesto pasa a ÁCIDO FUMÁRICO, mediante una OXIDACIÓN en la que se reduce el FADH a FADH2. El ácido fumárico pasa a ÁCIDO MÁLICO mediante la incorporación de una molécula de H2O. Y finalmente este pasa mediante una OXIDACIÓN a ácido oxalacético otra vez, reduciéndose en este proceso el NAD+ a NADH. De manera que por cada vuelta se obtienen 3NADH, 2FADH2 (que se van a la cadena de transporte electrónico para obtener ATP), 2CO2 (que se libera) y GTP (que es análogo al ATP). De esta manera se completa la degradación del esqueleto carbonado de la glucosa que poseía 6 carbonos, pues 2 de ellos han sido perdidos en la descarboxilación oxidativa, y como son 2 acetil-CoA los que tenemos se dan 2 vueltas al ciclo de krebs por lo que se desprenden 4 carbonos que sumados a los 2 de la fase anterior conforman los 6 C de la glucosa.

40. Metabolismo celular: -Define los conceptos de metabolismo, anabolismo y catabolismo. -¿Son reversibles los procesos anabólicos y catabólicos? Razone la respuesta. -El ciclo de Krebs es una encrucijada metabólica entre las rutas catabólicas y las rutas anabólicas ¿Por qué?
METABOLISMO: es el conjunto de reacciones químicas que tienen lugar en el interior de las células para satisfacer las necesidades de materia y energía de la misma. Todas estas reacciones se relacionan en unas rutas o vías metabólicas, en las que cada reacción interviene como mínimo una enzima, de manera que el producto de una reacción es el sustrato de la siguiente. Hay 2 tipos de rutas el anabolismo y el metabolismo. Y el objetivo del metabolismo es la obtención de energía, a través de la degradación de materia orgánica, la obtención de materia precursora sencilla necesaria para la síntesis de materia orgánica y por último la biosíntesis de moléculas orgánicas.
ANABOLISMO: es una vía constructiva del metabolismo, es decir, es la ruta en la que se produce la síntesis de moléculas complejas a partir de moléculas sencillas y energía.
CATABOLISMO: es un conjuntos de reacciones del metabolismo cuyo objetivo es la obtención de moléculas precursoras sencillas y energía, a partir de la degradación de biomoléculas.
Los procesos del anabolismo y el catabolismo son reversibles, ya que los productos de una de esas rutas metabólicas pueden ser los reactivos de la otra, aunque a veces las vías no son las mismas y se requiere de enzimas diferentes.
El ciclo de krebs es una encrucijada metabólica entre las rutas del anabolismo y el catabolismo ya que se puede llevar a cabo tanto en procesos catabólicos (B-oxidación) como en anabólicos (obtención de polisacáridos, trigliceridos,...)
41. Quimiosíntesis: Concepto e importancia biológica.
La quimiosíntesis consiste en la síntesis de ATP a partir de la energía desprendida en las reacciones de oxidación de sustancias inorgánicas, y el posterior uso de ese ATP para transformar la materia inorgánica en materia orgánica. Los organismos que llevan a cabo este proceso son los quimioautótrofos y son todos bacterias.
En esta se producen 2 fases: una primera en la que se oxidan sustancias inorgánicas, dando lugar al ATP por fosforilación oxidativa, además este se utiliza para provocar un transporte inverso de electrones en la cadena respiratoria para dar lugar a NADH. Y en la segunda el carbono se fija a partir del CO2 en el ciclo de calvin.
Muchos de los compuestos inorgánicos reducidos que se utilizan vienen de la descomposición de la materia orgánica que llevan a cabo los hongos y bacterias de la putrefacción. De esa manera la bacterias quimioautótrofas lo que hacen es oxidarlos transformándolos en sustancias minerales que pueden ser absorbidas por las plantas. Con lo cual los organismos quimiosintéticos desempeñan un papel imprescindible al cerrar los ciclos biogeoquímicos.
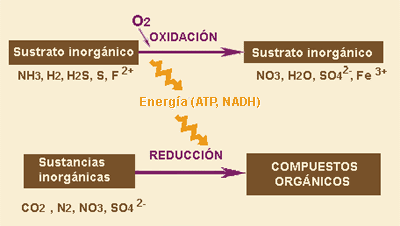
42. Importancia de los microorganismos en la industria. Fermentaciones en la preparación de alimentos y bebidas. Fermentaciones en la preparación de medicamentos.
Microorganismos como las levaduras o las bacterias son importantes en la industria ya que en ellos se produce un proceso que se conoce como fermentación, en la cual se obtienen una serie de productos que cuentan con un gran interés en el mundo de la industria y la alimentación.
La fermentación es un proceso catabólico en el que no hay cadena de transporte electrónico como hay en la respiración celular. De esa manera no hay tampoco hay ATP-SINTETASA, lo que explica porque siempre se produce una fosforilación a nivel de sustrato, lo que a su vez explica el escaso rendimiento energético. Además el último aceptor no es el oxígeno ya que la fermentación es un proceso anaerobio, sino que es una molécula orgánica.
Esta como ha sido mencionado anteriormente, se produce en microorganismos como las levaduras o las bacterias, pero también en las células musculares, cuando tras haberse realizado un ejercicio intenso no se ha producido la llegada de suficiente oxígeno por la sangre.
El verdadero significado de la fermentación está en la siguiente idea. Cuando una molécula de glucosa se degrada en la glucólisis da lugar a dos moléculas de NADH, pero ese NADH en la célula es limitado por lo que deberá reciclarse para que el proceso no sea parado. En su caso las células aerobias lo que hacen es reducir el NADH cediendo este sus electrones al oxígeno, que es el último aceptor de electrones. Mientras que en las células anaerobias, el NADH cede sus electrones a una molécula orgánica que puede ser el ácido pirúvico o algún producto de la degradación de este, mediante un o varias reacciones adicionales a la glucólisis. A esas reacciones adicionales se les llama fermentación y por tanto el principal objetivo de esta es el reciclaje del NADH.
La fermentación consiste en la transformación del ácido pirúvico en productos más sencillos que son diferentes en función del tipo de fermentación que se de, y es que existen varios tipos de fermentaciones que proporcionan una amplia gama de productos con intereses industriales y alimenticios.
Un tipo de fermentación es la homoláctica, que solo tiene una etapa en la que el ácido pirúvico acepta un par de electrones del NADH y se reduce a ácido láctico que es el producto final. Este tipo de fermentaciones se da en bacterias como el Lactobacillus, y se obtienen productos derivados de la leche, como queso, yogurt y requesón, de ahí el interés de esta fermentación en la industria alimenticia. Por otro lado encontramos la fermentación heteroláctico cuyo producto es mixto (ácido láctico, alcohol etílico y CO2).
Otro tipo de fermentación es la alcohólica, esta consta de 2 etapa, en la primera el ácido pirúvico pierde el grupo carboxilo en forma de CO2, convirtiéndose en acetaldehido y en la segunda el acetaldehido acepta un par de electrones del NADH transformándose en alcohol etílico que es el producto final. Pero existen aún más fermentaciones como la butírica y la putrida, en el caso de la butírica, se produce la descomposición de sustancias glucídicas de origen vegetal, almidón, celulosa,... en ácido butírico, CO2, hidrógeno y sustancias malolientes. Y en el caso de la putrida es la descomposición pero de sustancias de naturaleza proteica o aminoacídica, dando lugar a sustancias orgánicas y malolientes, algunos de los productos que son poco desagradables de esta fermentación se utilizan para producir los sabores típicos del queso, del vino,...
43. Fermentaciones y respiración celular. Significado biológico y diferencias.
La fermentación es un proceso catabólico en el que no hay cadena de transporte electrónico como hay en la respiración celular, y que consiste en la obtención de materia orgánica sencilla a partir de materia orgánica más compleja.
El verdadero significado de la fermentación está en la siguiente idea. Cuando una molécula de glucosa se degrada en la glucólisis da lugar a dos moléculas de NADH, pero ese NADH en la célula es limitado por lo que deberá reciclarse para que el proceso no sea parado. En su caso las células aerobias lo que hacen es reducir el NADH cediendo este sus electrones al oxígeno, que es el último aceptor de electrones. Mientras que en las células anaerobias, el NADH cede sus electrones a una molécula orgánica que puede ser el ácido pirúvico o algún producto de la degradación de este, mediante un o varias reacciones adicionales a la glucólisis. A esas reacciones adicionales se les llama fermentación y por tanto el principal objetivo de esta es el reciclaje del NADH.
Mientras que la respiración celular es el proceso de la degradación completa de la glucosa en presencia de oxígeno (camino que se sigue tras haberse realizado la glucólisis). Por tanto la finalidad del mismo es la degradación de la molécula de glucosa, para dar lugar a H2O, CO2 y energía en forma de ATP.
Con respecto a las diferencias son numerosas, en primer lugar en las fermentaciones no hay cadena de transporte de electrones pero en la respiración si, por otro y como consecuencia de lo anterior no hay ATP-SINTETASA en las fermentaciones por lo que la fosforilación que se produce siempre es a nivel de sustrato, esto explica el escaso rendimiento energético de esta (solo 2 ATP de la glucólisis) . Mientras que en la respiración si que hay ATP-SINTETASA, por lo que se produce fosforilación oxidativa, eso se traduce en un mayor rendimiento energético.
Además la fermentación es anaerobia por lo que no hay presencia de O2 y por tanto este no puede ser el último aceptor de electrones, de manera que lo será una molécula orgánica. Mientras que en la respiración si que hay O2, son reacciones aerobias, en las que este es el último aceptor de electrones.

44. A) En la figura se indican esquemáticamente las actividades más importantes de un cloroplasto. Indique los elementos de la figura representados por los números 1 a 8.
B) Indique mediante un esquema, qué nombre reciben las distintas estructuras del cloroplasto. ¿En cuál de esas estructuras tiene lugar el proceso por el que se forman los elementos 4 y 6 de la figura? ¿Dónde se produce el ciclo de Calvin?
C) ) Explique brevemente (no es necesario que utilice formulas) en qué consiste el ciclo de Calvin.
c) Es la fase oscura de la fotosíntesis en la que no se requiere de luz, en ella lo que ocurre es la síntesis de materia orgánica a partir de CO2, utilizándose el ATP y el NADPH obtenido en la fase luminosa. Esta fase se da lugar en el estroma de los cloroplastos. A grandes rasgos se podría decir que es la fijación del carbono a partir de CO2, para dar lugar a una molécula orgánica más estable. Este se divide en tres fases que son la fijación del CO2, la reducción del CO2 fijado y la regeneración de la ribulosa 1,5-difosfato, las cuales se llevan a cabo de la siguiente manera:
Primeramente la pentosa ribulosa 1,5-difosfato se une en el estroma al CO2 atmosférico gracias a la enzima ribulosa 1,5-difosfato carboxilasa oxidasa, conocida con el nombre de ribusco, que es la que posibilita esa fijación del CO2 en la pentosa. De esa manera se da lugar a un compuesto inestable de 6 átomos de carbono el cual se divide en 2 moléculas de ácido 3-fosfoglicérico, que son moléculas de 3 átomos de carbono.
Posteriormente, se produce la reducción de ese CO2 fijado. En este caso, primero el ácido 3-fosfoglicérico es fosforilado, consumiéndose de esa manera ATP (obtenido en la fase luminosa, que se reduce a ADP y Pi) y así obtenemos 1,3-difosfoglicérico. Y a continuación este se reduce consumiéndose el NADPH (obtenido en la fase luminosa que se reduce a NADP+) y así obtenemos gliceraldehido 3-fosfato. Este gliceraldehido-3-fosfato puede seguir dos vías, la mayor parte se invierte en regenerar la ribulosa1,5-difosfato y el resto para otras biosíntesis (puede quedarse en el estroma para la síntesis de almidón, ácidos grasos, aminoácidos,.. o puede ir al citosol para dar lugar a la glucosa o fructosa). En el caso de la regeneración de la ribulosa 1,5-difosfato, lo que ocurre es que el gliceraldehido 3-fosfato se transforma en ribulosa-5-fosfato mediante un proceso complejo en el que se suceden una serie de compuestos de 3,4,5 y 7 carbonos. Al final se regenera la ribulosa 1,5-difosfato mediante la fosforilación directa con ATP. (este proceso es conocido como ciclo de las pentosas fosfato)
Por tanto en el ciclo de calvin por cada CO2 que se incorpora se requiere de 2 NADPH y 3 ATP. Este es un proceso que no requiere de energía luminosa ni de clorofila, pero se suele llevar a cabo durante el día porque es cuando se sintetizan el ATP y el NADPH.

1.
CO2
2.
Ribusco (ribulosa 1,5 difosfato)
3.
ADP
4.
ATP
5.
NADP+
6.
NADPH
7.
H2O
8.
O2

45. A) la figura representa esquemáticamente las actividades más importantes de una mitocondria. Identifique las sustancias representadas por los números 1 a 6.
B) La utilización de la energía liberada por la hidrólisis de determinados enlaces del compuesto 4 hace posible que se lleven a cabo reacciones energéticamente desfavorables. Indique tres procesos celulares que necesiten el compuesto 4 para su realización.
-
B-oxidación de ácidos grasos.
-
Fotosíntesis.
-
Glucólisis.
C) En el esquema, el compuesto 2 se forma a partir del compuesto 1 , que a su vez, proviene de la glucosa. ¿Sabría indicar otra sustancia a partir de la cual se pueda originar el compuesto 2?
Otra manera de obtener acetil-CoA es la B-oxidación de los ácidos grasos o hélice de Lynen, en la que por cada vuelta el acil(graso)-CoA pierde en se oxidación un acetil-CoA.

1.
Ácido pirúvico
2.
Acetil-CoA
3.
ADP
4.
ATP
5.
NADH
6.
Oxígeno
46. a) El Esquema representa un cloroplasto ¿Qué denominación reciben los elementos indicados por los números 1-7?
b) En los cloroplastos, gracias a la luz, se producen ATP y NADPH. Indique esquemáticamente, como se desarrolla este proceso.
c) Las moléculas de ADN de los cloroplastos y las mitocondrias son mucho más pequeñas que las bacterias. ¿Contradice este hecho la hipótesis de la endosimbiosis sobre el origen de las células eucarióticas?
No, ya que realmente la endosimbiosis nos explica que el origen de la célula eucariótica viene a partir de una línea celular que fagocitó una bacteria con la que llevo a cabo una endosimbiosis, es decir la bacteria fagocitada no se digirió. En esta, la célula le proporcionaba materia a la bacteria, la cual se encargaría de oxidarla dando lugar a energía en forma de ATP para la célula. Así surgió la primera célula animal.
Mientras que por otro lado algunas de estas células fagocitarían cianobacterias, capaces de realizar la fotosíntesis, llevando a cabo también una relación endosimbiótica con estas lo cual permitiría realizar la fotosíntesis dentro de la otra célula, surgiendo así la primera célula vegetal.

1.
Espacio intermembrana
2.
Membrana plastidial interna
3.
Membrana plastidial externa
4.
Tilacoides de estroma
5.
ADN plastidial
6.
Estroma
7.
Tilacoides de grana
47. a) En el interior de este cloroplasto hay almidón. Explique, mediante un esquema, como se forma la glucosa que lo constituye.
b) Indique tres similitudes entre cloroplastos y mitocondrias.
Entre las similitudes existentes entre cloroplastos y mitocondrias encontramos las siguientes:
-
Ambos presentan una doble membrana.
-
Ambos se encuentran encuentran en células eucariotas.
-
Ambos tienen material genético propio. (ADN circular)
-
Ambos tienen cadena de transporte electrónico, en el caso de la mitocondria se encuentra en las crestas mitocondriales de la membrana mitocondrial interna. Y en el caso de los cloroplastos en la membrana de los tilacoides.


48. a) El esquema representa una mitocondria con diferentes detalles de su estructura. Identifique las estructuras numeradas 1 a 8.
b) Indique dos procesos de las células eucariotas que tengan lugar exclusivamente en las mitocondrias y para cada uno de ellos establezca una relación con una de las estructuras indicadas en el esquema.
-
Fosforilación oxidativa, la cual se produce en la cadena transportadora de electrones, que está en la crestas mitocondriales de la membrana mitocondrial interna.
-
Ciclo de krebs, el cual se produce en la matriz mitocondrial.
c) Las mitocondrias contienen ADN. Indique dos tipos de productos codificados por dicho ADN.
Proteínas mitocondriales y un número pequeño de polipéptidos.

1.
Matriz mitocondrial
2.
Crestas mitocondriales
3.
Mitorribosomas
4.
Membrana mitocondrial interna
5.
Membrana mitocondrial externa
6.
Espacio intermembrana
7.
ATP-SINTETASA
8.
Cadena transportadora de electrones
Los esquemas de los vídeos de la lesson plan se encuentran incluidos en los esquemas anteriores.
A continuación adjunto un pdf con las actividades del tema: